Bakterilerde Genetik Madde Aktarımı
Prof. Dr. Mustafa Arda
Ankara Üniversitesi Veteriner Fakültesi
Temel Mikrobiyoloji 1
01. Giriş
02. Transformasyon
03. Konjugasyon
04. Transdüksiyon
04.01. Generalize Transdüksiyon
04.02. Özel Transdüksiyon
04.03. Abortif Transdüksiyon
05. Genetik Rekombinasyonlar
05.01. Homolog (Genel) Rekombinasyon
05.02. Site Specific (Özel ) Recombination
05.03. Nonhomolog Rekombinasyon
06. Prionlar ve Replikasyonları
01. Giriş
Bakteriler arasında genetik materyalin (DNA segmentinin kısa veya uzun bir kısmı veya tümü) bir vericiden (donör) başka bir alıcıya (resipient) transferi, normal koşullar altında, in vivo veya in vitro olarak kolayca gerçekleşebilmektedir. Aktarılan genler, eğer bakteriler birbirlerine genetik olarak çok yakınsalar veya DNA sekansları arasında büyük bir homoloji varsa, alıcının kromozomu ile kolayca birleşebilmektedirler. Eğer, homoloji çok azsa veya ayrı cinslere ait türler ise bu birleşme çok daha az oranda olabilmektedir.
Burada iki konuyu birbirinden ayırmada yarar vardır. Yabancı DNA'nın vericiden alıcıya transfer mekanizması ile, hücre içine giren segmentlerin alıcının kromozomu ile entegrasyonu farklı olgulardır. Birbirinden ayrı düşünülmesi gerekir.
Vericinin DNA segmentlerinin alıcı bir bakteriye aktarılması ya doğal olarak ve fakat çok az sıklıkla 10-7 - 10-9) meydana gelebilir veya suni olarak özel teknikler kullanılarak (elektroporasyon, CaPO4 presipitasyon, mikroenjeksiyon, mikroprojektil, vektörler, vs) bu oran yükseltilebilir (10-3 - 10-4) gibi.
Genetik materyaller sadece bakterilere ve ökaryotiklere ait değildir. Plasmid, faj, virus, transpozon ve insersiyon sekansları da olabilir ve aktarılabilirler. Bunlara ait bilgiler kendilerine özgü bahislerde verilmiştir.
Gen transferleri sonunda, alıcı bakteriler, kendi kromozomuna integre olan yabancı DNA sekansında bulunan genin (veya genler) taşıdığı özel karakterler yönünden pozitif hale dönüşebilirler.
Bakteriler arasında oluşan bu doğal gen transferi yanı sıra, biyoteknolojik yöntemlerle de (rekombinant DNA teknolojisi) gen aktarımlarını başarı ile gerçekleştirmek olanak dahilindedir. Bu teknikte, bir vericide (prokaryotik ve ökaryotik) bulunan ve çok önemli bir proteinin sentezini kodlayan geni (genleri), restriksiyon endonuklease enzimleri ile keserek çıkarmak, bir aracı moleküle (vektör) bağlayarak oluşturulan bu rekombinant vektörü (plasmid, faj, cosmid, mekik vektörler, virus, bakteri) alıcı bir organizmaya (prokaryotik veya ökaryotik) transfer etmek ve burada genin ekspresyonunu sağlamak mümkündür (gen klonlaması, moleküler klonlama). Çok yararlar sağlayan bu teknoloji hastalıkların teşhis, kontrol ve sağaltımın da olduğu kadar araştırmalar da da çok büyük katkıları bulunmaktadır.
Bakteriler arasında doğal gen transferleri başlıca 3 mekanizma ile gerçekleşmektedir.
1) Transformasyon
2) Konjugasyon
3) Transdüksiyon
02. Transformasyon
Bir mikroorganizma, kendisine DNA kompozisyonu yönünden çok yakın olan, diğer bir mikroorganizmaya ait genetik materyal (DNA segmentleri) içeren bir ortamda üretilirse, belli bir süre sonra, bu sıvı ortamdan katı besi yerine ekimler yapılırsa, bazı kolonilerin değişik morfolojide olduğu ve bunların, genetik materyali veren ölü mikroorganizmanın orijinal kolonilerine benzediği görülür. Örn, D. pneumoniae 'nin II-R suşu, kendisini oluşturan II-S suşundan ekstre edilen DNA materyalini ihtiva eden uygun bir sıvı ortamda ve uygun bir süre temasa bırakıldıktan sonra, katı besi yerine yapılan ekimlerde bazı kolonilerin II-S karakterinde olduğu görülür.
Bu denemede, II-S suşu öldürülmüş ve elde edilen DNA ekstraktları ortama katılmıştır. Canlı olan II-R suşu ortamda bulunan II-S suşuna ait bazı genetik materyalleri almış ve bu elementlerde taşınan karakterler yönünden pozitif hale gelmiştir (II-S haline dönüşmüştür).
Transformasyonun genel şeması yandaki şekilde gösterilmektedir. | 
|
Yukarıda bildirilen in vitro transformasyon, aynı şekilde, in vivo olarak ta oluşturulabilir. İn vivo transformasyon, Griffith'in deneyi ile gösterilebilir. Farelere, önce virulent ve kapsüllü olan II-S suşundan mutasyonla elde edilen canlı II-R suşu (avirulent ve kapsülsüz) verilir. Bunun arkasından, ısı ile öldürülmüş olan II-S suşundan fazla miktarda şırınga edilir (aslında II-R suşu pneumonia oluşturmadığı gibi, II-S 'de öldürülmüş olduğu için infeksiyöz yeteneği yoktur). Ancak, deneme sonunda ölen farelerden katı besi yerine yapılan ekimlerde, üreyen koloniler arasında, bazılarının II-S karakterinde (kapsüllü ve virulent) oldukları görülür. Bu durum, aynı in vitro deneyde olduğu gibi, II-R suşları, fare vücudunda, II-S 'lere ait bazı genetik materyalleri almış ve II-S karakterine dönüşmüş ve fareleri öldürmüştür.
Bu iki (in vivo ve in vitro) denemede, ortak olan nokta, verici hücre (bakteri) DNA parçacıklarının alıcı hücre (bakteri) kromozomu ile birleşmiş olması ve verici hücreye ait karakterleri yönünden alıcıda pozitifliğin elde edilmesidir. Her iki hücrenin de antijenik yakınlığının çok fazla olması (birbirinden mutasyon ile türemiş olmaları) gen alışını kolaylaştırmaktadır. Antijenik yakınlığı olmayan bakteriler arasında transformasyon, genellikle, nadirdir.
Bir DNA segmentinin, diğer bir bakteri kromozomu ile birleşebilmesi için, her ikisinde de baz sıraları bakımından bir homologluğun olması şarttır. Bu nedenle, birleşebilen ve birbirinin komplementleri olabilen bu bölge ile DNA parçacıklarındaki A+ T/G + C oranının aynı veya birbirine çok yakın olması da gereklidir. Bu takdirde, verici hücre DNA segmenti, kendine homolog olan, alıcı hücre DNA'sının özel bölgesiyle ilişki kurar ve buradan birleşebilirler (integrasyon). Bazı durumlarda, verici hücre DNA'sı alıcı hücreye girse bile özel alıcı bölgeler (homolog yanlar) olmadığı sürece birleşme olamaz. Homolog rekombinasyonun meydana geldiği durumlarda, verici hücre DNA parçacığı, alıcının kromozomunun bir parçası veya devamı haline gelir. Bununla birlikte replike olur ve hücrenin bölünmesi sırasında da bir kopyayı diğer kardeş hücreye aktarılır.
Transformasyonda, alıcı ve verici (donör) hücrelerin DNA'larının homolog oluşları yanı sıra bazı önemli faktörlerin de etkisi bulunmaktadır. Uygun ortam ve optimal koşullarda, üreyen hücrelerden ancak 10-7-10-8 kadarının, DNA segmenti alma yeteneği vardır. Bu nedenle üreme sırasında bazı hücrelerin belli bir fizyolojik duruma ulaşmasının ve DNA parçasını alabilecek kabiliyete (permeabiliteye) kavuşmasının da önemi fazladır. Diğer bir deyimle alıcı hücrenin diğerlerinden farklı ve kompetent olması gereklidir. Diğer önemli noktalardan biri de, DNA segmentlerinin, 0.3-8 x 105 dalton molekül ağırlığından aşağı olmaması durumudur. Çift iplikçikli olmayan ve yukarıda bildirilen ölçülerden küçük olanlar alıcı hücrelere kolayca giremezler.
Alıcı hücrenin yüzeyi, verici hücrenin DNA parçacığının adsorpsiyonu için önemlidir. Alıcı hücre yüzeyinde özel ve belli alıcı bölgeler (reseptörler) bulunmaktadır. Kapsüllü bakterilerde adsorpsiyon ve içeri giriş olgusu çok zordur. Yapılan çalışmalar, bazı bakterilerde 75 kadar böyle özel alıcı adsorpsiyon bölgesinin bulunduğunu ortaya koymuştur. Kapsülün bulunmadığı ve diğer koşulların uygun olduğu durumlarda, adsorpsiyon gerçekleşebilir. Hücre yüzeyine adsorbe edilen DNA segmenti, eğer içeri girme fırsatını bulamaz veya girmede gecikirse tuz, pH ve deoksiribonuklease (endonuklease) tarafından etkisiz hale getirilir veya lize edilir. Bu etkilerden kurtulabilenler içeri girerler (penetrasyon). Yukarıda bildirilen engeller, bakteriler için bir korunma veya bir sınırlama mekanizmasıdır. Verici hücre DNA'sının çift iplikçikli olması rekombinasyon bakımından önemlidir. Ancak, bazen alıcı hücre DNA'sı ile sadece bir iplikçik birleşir, diğeri endonuklease tarafından lize edilir. Birleşen iplikçiğin karşısında, alıcı hücre, hemen komplementerini sentezler. Böylece birleşme bölgesi çift iplikçik haline dönüştürülür (rekombinasyon).
DNA segmenti alıcı hücre yüzeyindeki özel bölgelere adsorbe olduktan ve içeri girdikten sonra, bakteri türleri arasında değişiklik göstermek üzere, 10 dakika ile birkaç saat süre içinde kompetent hücre DNA'sı ile birleşme olur. Aradan geçen bu süreye eklips (latent) dönem adı verilir. Hücreye giren DNA segmenti çift iplikçik halinde kalabilir ve sirküler forma dönüştürüldükten sonra alıcının DNA'sı ile de birleşebilir. Ekzogenotun alıcı hücre DNA'sındaki özel komplementer bölgeyle yan yana gelmesi gereklidir. Buda sağlandıktan sonra, ekzogenot ile endogenot arasında homolog rekombinasyon gerçekleştirilir. Oluşan hibrid DNA'nın uçları ligase enzimi aracılığıyla birleştirilerek rekombinasyon tamamlanır. Böylece integre olan ekzogenot alıcı hücrenin DNA'sının bir parçası ve devamı haline gelir ve birlikte replike olur. Ancak, alınan karakterlerin yeni nesillerde görülebilmesi için 2-3 generasyonun geçmesi gereklidir.
Adsorpsiyon olayı geriye dönüştürülebilir. Eğer, alıcı hücre yüzeyindeki reseptörler (alıcı bölgeler) tripsinle veya DNA parçacıkları DNase ile muamele edilirse adsorpsiyon gerçekleşemez. İçeri girdikten ve integrasyon olduktan sonra artık DNase etkilemez. Eklips dönemi bakteriler arasında değişiktir. H. influenzae de bu süre çok kısadır. Bazı bakterilerde ise birkaç saat uzayabilir.
D. pneumoniae 'den başka H. influenzae, stafilokok, bitki patojenleri, basiller, nitrojeni fikse eden mikroorganizmalar arasında da, transformasyon tespit edilmiştir. Enterobakteriler arasında transformasyon pek başarılı olmamakla beraber, son yıllarda, yüksek kalsiyum iyonları konsantrasyonlarında E. coli 'de de transformasyon saptanmış olduğu bildirilmektedir.
Transformasyonla, verici bakterilere bazı özel karakterler aktarılabilmiştir. Bunlar arasında laktoz ve galaktoz pozitif genler, antibakteriyel maddelere karşı dirençlik, virulens, v.s. sayılabilir. Bu faktörler alıcının DNA'sı ile birleştikten sonra, transkripte olarak mRNA'ya aktarılır ve buradan da transle edilerek, hücrede yeni karakterlerin ortaya çıkmasına neden olurlar.
Transformasyondan yararlanarak bakterilerin kromozom haritaları çıkarılabilir. Bu teknikle elde edilen haritaların yardımıyla gen sıraları, genlerin birbirine uzaklıkları ve birbirlerine olan ilişki durumları öğrenilebilir. Ancak, en iyi haritalar konjugasyon ve transdüksiyon ile yapılmaktadır.
Transfeksiyon: Virus, plasmid, faj, bakterilerin ve DNA segmentlerinin kompetent hücrelere özel yöntemlerle aktarılması olayına transfeksiyon adı verilir. Örn, bakteride sentez edilen fajlardan bazıları kapsid taşımayabilir veya faj DNA'sı sentezlendikten sonra kapsidle birleşmeyebilir. Bakteri lize olduktan sonra serbest kalan bu fajlar ortamda hazır bulunan kompetent hücreler tarafından alınabilirler. Ancak, bu olay çok sınırlı ve nadirdir. E. coli K12'de ve sferoplastlarda transfeksiyona rastlandığı bildirilmiştir.
Hücrelere DNA segmentinin fazlaca girmesini kolaylaştırmak için, CaPO4'la presipitasyon, mikroenjeksiyon, elektroporasyon, vs özel tekniklerden yararlanılır.
03. Konjugasyon
Konjugasyon, verici hücre DNA'sının tümünün veya bir segmentinin, ya bu iki hücrenin direkt teması veya seks pilusları aracılığı ile alıcıya aktarılması olayına verilen bir isimdir. Bakterilerde konjugasyon olgusunu aydınlatan ilk çalışmalar, J. Lederberg ve E.L. Tatum'un (1946) araştırmalarından kaynaklanmaktadır. Araştırıcılar, iki amino asit ekzotrofik karakteri taşıyan 2 E. coli K-12 mutantı üzerinde yaptıkları çalışmalar ile konjugasyona açıklık kazandırmışlardır. Bu denemede, biri, amino asit A- ve B- için okzotrofik, fakat C+ ve D+ amino asitleri sentezleyebilen (A-B-C+D+), diğeri ise bunun tersi olan (A+B+C-D-) iki mutant suş bir besi yerinde karıştırarak bir süre (6-7 saat) inkubasyonda tuttuktan sonra (bu aminoasitleri içermeyen) minimal besi yerine ekilmiştir. Bu iki suş ayrı ayrı bu besi yerlerinde ürememelerine karşın, birlikte olduktan sonra ekildiklerinde bazı kolonilerin meydana geldiği ve bu kolonilerdeki E. coli 'ler incelendiğinde 4 amino asit yönünden de (A+B+C+D+) pozitif oldukları görülmüştür. Ancak bu olgu 10-6 kadardır.
Konjugasyonun genel şeması yandaki şekilde gösterilmektedir. | 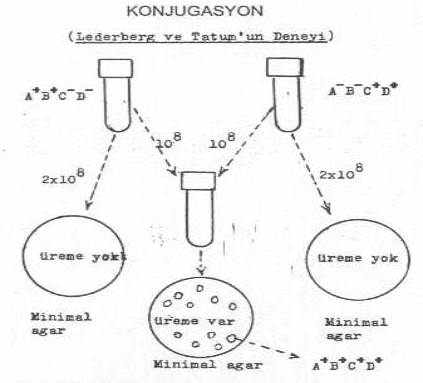
|
B.Davis (1950) U-tüpü kullanılarak yaptığı bir çalışmada konjugasyon için direkt temasın gerekli olduğunu ortaya koymuştur. U-tüpünün alt kısmına yerleştirilen ve bakteri tutan bir filtre ile sadece besi yerinin birbirine geçişi sağlanmasına karşın bakterilerin geçişi önlenmiştir. Tüpün içine minimal besi yeri konduktan sonra, bir yarımına met+, thr,- leu-, thi- ve diğer yarımına da met-, thr+, leu+, thi+ E. coli mutant suşları ekilmiş ve 4 saat inkubasyonda tutulduktan sonra minimal besi yerlerine ekildiğinde hiçbir koloninin oluşmadığı gözlemlenmiştir. Bu durum gen aktarımda direkt temasın önemli olduğu ortaya koymaktadır.
Lederberg ve Tatum'un denemelerinde direkt temas üzerinde durmamışlardır. Davis, bu çalışması ile direkt temasın konjugasyonda çok önemli faktör olduğunu ortaya koymuştur.
Davis ’in U Tüp Deneyi yandaki şekilde gösterilmektedir. | 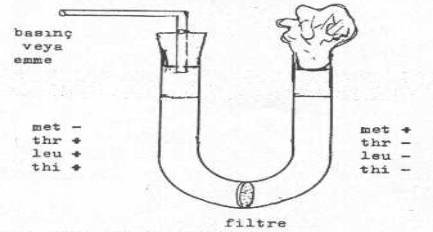
|
Protoplast füsyon tekniği de (bakteri, basil, streptomyces, maya, bitki, vs.) gen aktarımı için kullanılmaktadır.
İki ayrı türe ait protoplastlar elde edildikten sonra, bunlar, PEG'lu ortamda direkt temasa getirilirler. Uygun bir süre ve ısıda tutulan protoplastlar arasında birleşme ve gen aktarmaları kolayca olabilmektedir.
Maya hücrelerinde protoplast füzyonu yandaki şekilde gösterilmektedir. | 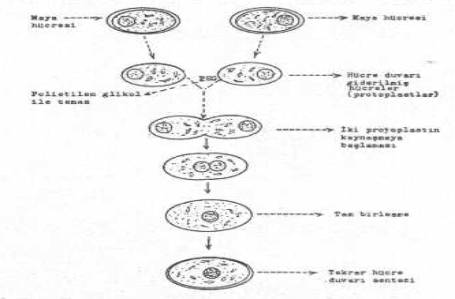
|
Konjugasyon fenomeninde, verici durumda olma kabiliyetini, genellikle, hücrede bulunan ve nakledilebilir bir genetik element olan plasmidler tayin ederler. Bu plasmide fertilite faktörü (F-faktörü veya seks faktörü) adı verilir. Bu faktörü alan hücreler taşınan karakterler yönünden pozitif hale gelirler. Aktarma olayı 10-5-10-6 oranında meydana gelmektedir.
İki hücre arasında konjugasyonun gerçekleşebilmesi için, bunların birbirleriyle temasa gelmesi gerekir. Bu olguyu, direkt temas kadar, genellikle, verici hücrelerdeki seks pilusları da yaparlar. Seks piluslarının sentezlenmesini hücre içinde bulunan seks faktöründeki özel genler sağlarlar. Bunlar, diğer normal piluslardan (fimbria) daha uzun ve kalındırlar. Ortaları boş olduğundan bir boru ve geçit köprüsü görevini yaparlar. Genetik materyal buradan geçerek alıcıya aktarılır. Normal pilusların gen aktarmasında rolleri yoktur.
Konjugasyonla aktarılabilen birçok ekstra kromozomal genetik element bulunmaktadır. Bunlar arasında başlıca, virulens faktörleri, fertilite faktörü (F-faktörü), dirençlilik faktörleri (R-faktörleri) ve kolisin faktörü (Col-faktörü) en önemlileri arasındadır.
F faktörü ihtiva eden hücrelerde bu faktörün kromozom üzerinde olması çok önemlidir. Ancak, böyle hücreler (Hfr, high frequency recombination) yüksek sıklıkta genetik materyallerini aktarabilirler. Kendisinde F faktörü olan, ancak bunu kromozomu üzerinde taşımayan hücrelerde (F+) genetik materyali aktarma oranı daha düşüktür.
Bir bakteride bulunan F faktörü, yalnız kendini değil aynı zamanda içinde bulunduğu hücrenin DNA'sının da aktarılmasını stimule eder. Bu nedenle, F faktörünün girdiği hücre, bu faktörü aktarmak için hemen reorganizasyona geçer ve seks pilusu sentezlemeye başlar. Normal piluslardan daha kalın, büyük ve ortası boş olan seks piluslarının sentezi kısa bir zamanda tamamlanır. Bu piluslar mekanik olarak giderilirse konjugasyon gerçekleşemez. Kopan veya koparılan seks pilusları kısa bir süre içinde (20-30 dakika) tekrar sentezlenirler. Bu sentez mekanizmasını, seks faktöründe bulunan özel genler yönetirler. Bu piluslar 7 nm. çapında olup aynen bir bakteriyofaj kuyruğu gibi görev yaparlar. Molekül ağırlığı 1200 kadar olan ve tek bir protein molekülünden oluşan bu piluslar, ancak elektron mikroskopla veya özel antikor teknikleriyle ortaya konabilirler.
F faktörü sirküler, çift iplikçikli DNA segmenti olup (plasmid) moleküler ağırlığı 35 x 106 ve uzunluğu da 32 nm'dir. Bu uzunluk, ortalama bir bakteri DNA'sının %1.2'si kadardır ve takriben 40-60 proteini kodlayabilir. F faktörü sadece bir transfer faktörü olup, diğer faktörleri (R, Col, v.s.) taşımaz ve kendi varlığını koruyabilecek ve aktarabilecek genetik bilgilere sahiptir.
F faktörünün stimulasyonu sonu oluşan seks piluslarına özel fajlar adsorbe olurlar. Bu fajlar başlıca iki tür olup biri küçük, yuvarlak, RNA yapısında (QB ve R17), diğeri ise flamentöz ve DNA karakterindedir. Bunlardan ilki seks pilusunun yüzeyine, ikincisi ise ucuna adsorbe olur. Bunlar bazen de piluslardan içeri girerler ve pilusun kanalını tıkayarak konjugasyona mani olabilirler. Ancak, bu fajlar DNA'larını pilustan içeri enjekte etmezler. Seks pilusu sadece bir köprü görevi yapmaz, aynı zamanda, F- hücreye bağlanmada da rol alır. Fakat, yapılan çalışmalar, seks pilusu olmadan da iki bakterinin yan yana gelmesi ve direkt teması hallerinde gen aktarmasının mümkün olduğunu ortaya koymuştur.
Laboratuvarlarda, konjugasyon oluşturmak için, F+ hücrelerle F- hücreler uygun bir inkübasyon ısısında belli bir süre aynı yerde bir arada bulundurulurlar. Her şeyin normal gittiği durumlarda transfer, mikroorganizmanın türüne göre değişmek üzere, konjugasyon 90-140 dakikada gerçekleşir. Köprüye en yakın bulunan genler en erken köprüye girerek alıcı bakteriye geçerler.
Bir dakika içinde takriben kromozomun %1'lik segmenti geçebilir ki bu da tüm kromozomun 11 nm'lik bir parçasını (veya iplikçik başına 5x106 nukleotid) oluşturur. Bu genetik uzunluk yaklaşık 20 rekombinasyon ünitesine tekâbül eder.
Aktarma sırasında Hfr hücrelerde, F faktörü, hücre DNA'sı ile birleşmiş olduğu yerden kopmakta ve bu faktör bakteri kromozomunun arka ucunda kalmaktadır. Bu durum, konakçı DNA'sının aktarılmasında itici bir güç yaratmaktadır. Sonda bulunması nedeniyle, eğer yeterli süre tanınmazsa veya kültürler çalkalanırsa, F faktörü alıcı hücreye aktarılamaz. Bu nedenle, Hfr x F- hücre birleşmelerinde genellikle F- hücreler elde edilir. Buna karşılık F+ x F- hücre birleşmesinde F+ hücreler daha fazladır. Halbuki, Hfr hücreler, F+'lere oranla, 1000 defa daha fazla konjugasyon yapma yeteneğine sahiptirler. Hfr hücrelerden F- faktörü alıcıya az oranda geçmesine karşın, donör DNA segmentinin geçme olanağı çok fazla olduğundan, gen aktarması yüksek sıklıkta gerçekleşebilmektedir.
Sirküler olan F faktörünün, alıcı hücre kromozomu ile birleşebilmesi için birbirlerinde homolog bölgelerin bulunması gereklidir. Birleşmeden önce sirküler olan F faktörü bakteri kromozomu üzerindeki homolog bölge yanına gelir. Birleşme homolog bölgeler arasında tek bir krosoverle gerçekleştirilir. Böylece, Hfr hücreler meydana gelirler. E. coli 'nin kromozomu üzerinde F- faktörü için homolog olan birçok bölge (8-10 kadar) bulunmaktadır. F faktörü bu bölgelerin bir tanesi ile integre olur. Hfr hücre oluştuktan sonra, bu faktör hücreyi seks pilusu oluşumu için tembih eder ve sentez mekanizmasını aktive eder.
Konjugasyon transferinden yararlanılarak bakteri kromozomunun gen haritası çıkarılmaktadır. Örn, Hfr x F- hücreler birbirleriyle temasa getirildikten sonra belli aralıklarla (her dakika veya iki dakikada bir) karışım çalkalanılarak gen transferine mani olunur veya kromozom koparılır.
Buradan yapılan ekimlerden meydana gelen koloniler aktarılabilen karakterler yönünden incelenirler. Böylece, dakika dakika, geçen kromozom segmentleri üzerindeki genlerin türü, sırası, birbirine olan ilişkisi ve uzaklığı saptanır. Yapılan çalışmalarda Hfr x P46 hücreleri ilk 5 dakika içinde prolin (pro+) sentezini idare eden genleri transfer etmekte, bunu 12 dakika sonra threonin (thr+) izlemektedir. Bu ilk iki gen arasında 7 dakikalık bir zaman dilimi bulunmaktadır. Diğer bir Hfr-H suşu ise, threonin genini ilk 8 dakika içinde aktarmakta, bundan 15 dakika sonra da prolin (pro+) geni geçmektedir. Bu son mikroorganizmada da iki gen arasında 7 dakikalık bir zaman dilimi vardır. Bu iki deneme transfer hızının sabit olduğunu kanıtlamaktadır. Bütün kromozomun aktarılması için geçen süre, mikroorganizma türüne göre değişmekte ve E. coli 'de 90, salmonellalar da ise 110-140 dakika arasındadır. E. coli üzerinde 3x 106 nukleotid çifti bulunmakta ve her dakikaya 3.3x104 nukleotid çifti veya yaklaşık olarak 33 gen isabet etmektedir. Kromozom daima 5'-ucundan alıcı bakteriye geçer ve bir kopyası daima verici hücrede kalır. Hfr x F- hücre birleşmesinde ilk geçen faktörler sırasıyla thr, leu, pro, lac, gal ve bunun arkasından ve daha yavaş olarak, his, ser, str, met geçerler. Ancak, her Hfr hücrenin kendine özgü transfer özelliği vardır ve bu durum DNA'daki kopma yerine göre değişir. Her Hfr'de kopma yeri farklı olduğundan aktarılan gen sırası da değişebilir. Ancak, çoğu zaman F faktörü kromozomun sonunda olacak şekilde kopma meydana gelir.
Yukarıdaki örneklerde de görüldüğü üzere, farklı Hfr suşlarında genlerin sırasında değişiklik bulunmaktadır. Birinde ilk gen prolin iken, diğerinde threonin olabilmektedir. Genler arasındaki zaman diliminin aynı olması bunların kromozom üzerinde aynı mesafelerde bulunduğunu gösterir. Ancak, transfer noktaları farklılık göstermektedir.
E. coli K 12 suşu, enterobakterilerin diğer türleriyle de (shigella, salmonella, serratia, proteus, v.s.) konjugasyon yapabildiği saptanmıştır. Ancak, bu ayrı cinslerle olan konjugasyon, E. coli türleri arasındakinden 1000 defa daha az sıklıkta olmaktadır. Çünkü, E. coli K 12 ile, S. typhimurium 'un bazı suşları arasındaki homologluk oranı %45, proteusla %6 ve S. flexneri ile %85 kadardır. Aralarında az çok homologluk durumunun bulunması nedeniyle konjugasyon olabilmekte ise de, sonunda integrasyonun olacağı söylenemez. Böyle homolog yanları az olan hücrelerde, F faktörü hücre içinde serbest halde bulunur (plasmid).
Hfr hücreler geri bir mutasyonla F+ hale gelebilirler. Böyle durumlarda, kromozomla birleşmiş ve onun bir devamı haline gelmiş bulunan F+ faktörü, kromozomdan ayrılarak sitoplazmada serbest hale geçer ve hücre de, bu nedenle, Hfr durumundan F+ hale gelmiş olur. Bu durumu ile F faktörü, plasmide benzer. Hücre içinde bağımsız olarak bulunan bu faktör, hücre DNA'sı ile birlikte ve aynı anda (senkronize) replike olur. Aynı zamanda, sitoplasmik membranda bulunan özel replikasyon bölgesini (mesosom) birlikte kullanırlar. Plasmid haline dönüşen F faktörü kendi varlığını devam ettirebilecek genetik bilgilere sahiptir. Hfr hücrelerinin F haline dönüşü (Hfr - F+) 10-5-10-6 oranında meydana gelir.
Bazı hallerde F faktörü, konakçı DNA'sı üzerinden ayrılırken, DNA'da yapıştığı bölgeden, konakçının bazı DNA parçacıklarını da alabilir. Bunlar, F faktörünün bir ucunda veya iki ucunda bulunabilirler. Böyle yabancı kromozom parçası ihtiva eden F faktörüne F'-prime faktör (F') ve alıcı hücreye de F' genot adı verilir. F' faktörünü taşıyan hücreler (F' prime), eğer çok önemli biyokimyasal olayları idare eden genleri kaybetmişse hücrenin ölümüne neden olabilir. Eğer, F faktörü, hücre DNA'sında iken galaktoz ve laktoz genlerinden birinin yanında ise, ayrılırken bu genlerden birini beraberinde alarak ekstra kromozomal genetik element haline geçebilir ve sitoplazmada F+-gal+ veya F+ - lak+ durumunda bulunabilir. Eğer bu F+ -gal+ veya F+ - lak+ plasmid başka hücreye (F¯ - gal- veya F-- - lak) transfer edildiğinde o hücreleri gal+ veya lak+ hale dönüştürürler. Böylece, oluşan hücreler sekonder F'-hücreler ve bu olaya da seksdüksiyon adı verilir. Eğer, sekonder F' hücreler, F- hücrelerle birleştirilirse iki tip transfer meydana gelebilir. Bazı hücreler sadece F' genotu, diğer kısmı ise, hem F' genot DNA'yı ve hem de bitişik kromozom DNA'yı transfer edebilirler. Bu ikinci tipte F integre olmuştur.
Bazı durumlarda da F-faktörü konakçı hücreyi terk ederek ayrılabilir ve böylece hücre F¯ hale gelir (Hfr ® F+ ® F¯). Diğer bir ifade ile hücreler, plasmidden kurtulabilirler (iyileşme). Bu olay, eğer F+ hücreler 42°C 'de ve durucu pozisyonda üretilirse gerçekleştirilebilir. Bazı durumlarda da akridin boyaları, hücre içinde bağımsız olarak bulunan F faktörünün replikasyonuna mani olmak suretiyle hücreleri F faktörünün etkisinden kurtarabilir. Bazı olaylarda da Hfr hücrelerinden F faktörü ayrılırken, F faktöründen bir kısmı alıcı hücre DNA'sında da kalabilir.
Yukarıdaki bilgiler ışığı altında F faktörü bir hücrede başlıca 3 pozisyonda bulunabilmektedir. Bunlar da:
1- Bağımsız genetik element (F+), (Plasmid)
2- Konakçı DNA'sı ile birleşmiş olarak (Hfr) (episom)
3- Bağımsız fakat, konakçı DNA'sından bir parça olarak (F' prime).
Bu durumları ile F faktörü, E. coli ‘nin lambda (l) fajına benzemektedir. Bu faj serbest veya virulent faj halinde bulunabileceği (plasmid) gibi, konakçı DNA'sı ile birleşmiş (profaj) (episom) veya konakçıdan laktoz genini almış olarak ta bulunabilir (F'prime).
04. Transdüksiyon
Genetik materyalin verici bir bakteriden alıcı bir bakteriye fajlar aracılığıyla nakledilmesine transdüksiyon adı verilir. Transdüksiyon suretiyle gen aktarılma olayına Gram negatif (Salmonella, E. coli, Shigella, Proteus, Vibrio, P. aeruginosa, vs.) ve Gram pozitif mikroorganizmalarda (Stafilokok ve basiller) rastlanılmıştır.
Fajlar, bakterileri parçalayan veya lize eden viruslardır (bakteriyofaj). Konakçıya özel olduklarından, bakteri fajları arasında da tür spesifitesi vardır.
Bazı fajların genomu, konakçı hücresine girdikten sonra orada replike olur ve bakteriyi parçalar (virulent veya vegetatif fajlar). Bir kısmı ise, infekte ettiği hücreyi lize etmeyebilir (temperate fajlar). Böyle fajlardan bazıları sitoplazmada kalır veya bazıları da konakçı DNA'sı ile birleşebilir. Böylece onun bir devamı haline gelebilir (profaj). İçerisinde profaj halinde bakteriyofaj bulunduran lizojenik hücreler, ultraviolet ışınlarına maruz bırakılırsa, profaj aktive olur ve konakçısını lize edebilir. Profajlar, yerleştikleri konakçı DNA'sından bir veya birkaç gen içeren bir segment parçasını alabilir ve bunu, adsorbe olduğu başka alıcı hücreye aktarabilir.
Yandaki şekilde A ve B bir bakterideki normal profajın replikasyonu, C ve D ise profajın bakteri DNA’sından bir segment olarak ayrılmasını göstermektedir. | 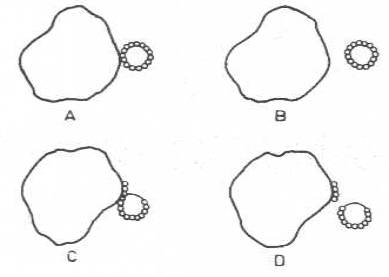
|
Transdüksiyon başlıca üç tarzda gerçekleştirilebilmektedir.
1- Generalize transdüksiyon,
2- Özel transdüksiyon,
3- Abortif transdüksiyon.
04.01. Generalize Transdüksiyon
Generalize transdüksiyonda, bakteri hücresi içinde faj olgunlaşırken, parçalanan konakçı DNA'sından bir segment, aynı yerde sentezlenen faj kapsidi içine tesadüfen girebilir. Böylece bu kapsid (fajın baş kısmı içinde), faj DNA'sından ayrı olarak ve değişik uzunlukta (50-100 gen içerebilir) bakteri DNA'sını da ihtiva etmiş olur. Böyle bir faj etrafa yayıldıktan sonra, başka bir bakteriyi infekte ettiğinde, kendi DNA'sı ile birlikte konakçı DNA'sını da alıcı hücreye aktarır. Eğer, faj, taşıdığı bu bakteri DNA'sı ile birlikte konakçı hücrenin kromozomu ile birleşirse, taşıdığı karakterler yönünden alıcı hücreyi pozitif hale getirir. Birkaç krossover dan sonra bakteri DNA'sı ile birleşebilen bu faj + bakteri DNA segmenti, alıcı hücre kromozomunun bir devamı haline gelir, birlikte replike ve transkripte olur ve taşıdığı genlerdeki bilgiler aynen mRNA'ya aktarılarak translasyona iştirak eder.
Generalize transdüksiyonda, bu görevi yapan fajlar, konakçı DNA'sının herhangi bir geninin veya belli bir segmentinin yanına yapışmazlar. Konakçı DNA parçalandığı zaman hücre içinde dağınık bir halde bulunan DNA parçacıklarından herhangi birinin, faj kapsidi içine tesadüfen girmesi ve içinde kalması sonu bu tarzda aktarılma şansına sahip olabilmektedirler. Bu nedenle her bakteri DNA segmentinin veya bunun üzerinde bulunan özel karakterlerin (genlerin) taşınma veya alınma olasılığı bütün genler için eşit olmaktadır.
Generalize transdüksiyon yapabilen fajlar arasında E. coli K 12'nin P1 fajı (M.A. 60 x 106) ve Salmonellalarda PLT 22 fajı (M.A. 26 x 106) vardır. Bu fajlarla gerçekleştirilebilen generalize transdüksiyon 10-5-10-6 oranındadır. E. coli K 12-P1 fajı konakçı DNA'sı ile birleşmez ve bir plasmid gibi hücrede bağımsız kalır ve müstakil olarak replike olur. Bu faj aracılığı ile, E. coli K 12 suşunun triptofan sentezinde görev yapan 5 genin sıralanması saptanabilmiştir. Salmonellalardaki PLT fajının hücreye adsorbe olabilmesi için konakçının somatik O-antijen 12 fraksiyonuna sahip olması gerekir. B. subtilis 'te bulunan SPIO ve PB 51 fajları da aynı şekilde generalize transdüksiyon yapabilmektedirler. Ancak, bu fajlar konakçı genomu ile integre olabilirler.
04.02. Özel Transdüksiyon
Özel transdüksiyon oluşturan fajlar konakçı DNA'sında belli bazı genlerin yanına yerleşirler ve hücre DNA'sı ile birleşerek profaj haline dönüşürler. Bu bakımdan da, episom karakteri gösteren F-faktörüne benzerler (Hfr). Konakçı hücrenin kromozomu ile birleşmiş bulunan faj, konakçı hücre UV-ışınlarına maruz bırakılırsa, aktive olur ve DNA'dan veya yerleştiği yerden ayrılarak konakçısını lize ederler. DNA'dan ayrılırken, yanında bulunduğu veya yanına yapıştığı bakteriye ait geni de birlikte alarak ayrılırlar. Böyle bir faj başka bir bakteri kültürüne inokule edilirse buradaki hücreleri infekte ettiğinde bu taşıdığı geni o bakteriye aktarır ve bakteriyi, gende bulunan özel karakterler yönünden, pozitif hale getirir.
Özel transdüksiyon yapan fajlar arasında E. coli K 12 'nin lâmbda fajı ve Ø 80 fajı örnek olarak gösterilebilir. Lâmbda fajı, E. coli 'de genellikle biotin ile galaktoz geninin arasına girer ve burada profaj olarak bulunur. Hücreden ayrılırken galaktoz genini de (gal+) birlikte alır. Eğer, bu faj, gal- bir mikroorganizmayı infekte ederse, bunu gal+ haline sokar. Bu tarz transdüksiyon 10-6-10-7 oranında gerçekleşmektedir. Triptofan genin yanına yerleşen Ø 80 fajı da; aynı lâmbda fajı gibi hareket eder.
Bazı durumlarda, profaj, konakçı hücresinden ayrılırken ve konakçı DNA'sından bir segment alırken buna karşılık kendi DNA'sından da bir kısmını, konakçı DNA'sı üzerinde bırakabilir. Böylece DNA'sında eksik genler bulunan faj, bu durumuyla başka bir bakteriyi infekte ettiğinde, bu alıcı hücre içinde eksik genlerin oluşturacağı veya sentezlediği önemli ürünleri bulamazsa ve bu bakteri de, eğer bu ürünleri sentezleyecek bilgilere haiz başka bir fajla infekte olmazsa, eksik olan faj genomu replike olamaz ve olgunlaşamaz. Böyle transdüksiyona da sınırlı transdüksiyon adı verilir.
04.03. Abortif Transdüksiyon
Fajlar tarafından, konakçısından alınan ve bazı karakterleri (genleri) taşıyan DNA segmenti, faj başka hücreyi infekte ettiğinde, bu hücreye aktarılır. Ancak, bu DNA parçası hücre içinde kalır, bakteri DNA'sı ile birleşmez. Bağımsız olarak hücre içinde bulunmasına karşın, hücre DNA'sı ile eş zamanda replike olma kabiliyetine de sahip değildir. Ancak, taşıdığı karakterler yönünden hücreyi pozitif hale getirebilir. Bakterinin her bölünmesinde, bu ekzogenot bir kardeş hücrede kalır ve onu pozitif hale sokar. Böylece DNA segmenti replike olmadan, bakterinin her bölünmesinde bir hücre kalmış olur.
İlk başlangıçta, bu ekzogenotu taşıyan ve fakat sonra, hücre bölünmesi sonu, diğer hücrede kalması nedeniyle sonradan bunu kaybeden bir bakterideki enzimler, müteakip bölünmeler de giderek azalır ve kaybolur. O zaman hücreler, bu enzimleri ihtiva etmediğinden negatif hale dönüşürler.
Abortif transdüksiyon yandaki şekilde gösterilmektedir. | 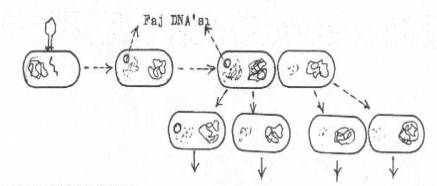
|
Yandaki şekilde genetik maddenin aktarım şekilleri gösterilmektedir. | 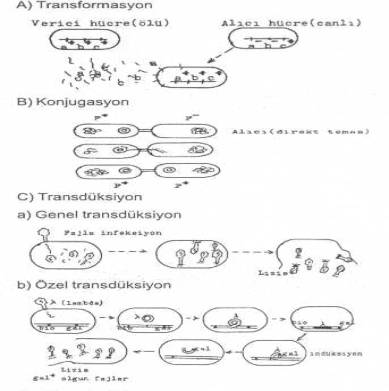
|
05. Genetik Rekombinasyonlar
Verici bir bakteriden, çeşitli tarzlarda aktarılan genetik materyallerin (ekzogenot) beklenilen etkinlikte olabilmesi için, sitoplazma içinde bulunmasından ziyade, alıcının kromozomu ile birleşmesi (integre olması) gereklidir. Bu olguya rekombinasyon adı verilmektedir. Bazı durumlarda alıcının sitoplazmasına giren yabancı DNA sekansları, burada kalabilir ve kromozomla birleşmeyebilir. Böyle durumlar, daha ziyade alıcı ile verici hücre DNA kompozisyonlarının farklı olduğu hallerde meydana gelebilir. Böyle olgular, sekanslar arasında homolog bölgelerin veya özel birleşme yerlerinin çok az veya olmamasından kaynaklanır.
Alıcının DNA'sına integre olan verici DNA sekansları, kromozomun bir devamı veya bir parçası haline gelerek, onunla birlikte ve eş zamanlı (senkronize) olarak replike olur ve ekspere edilir. Replike olan DNA segmenti, kardeş hücrelere de taksim edilir.
Eğer aktarılan yabancı DNA sekansında, alıcıda bulunmayan özel genler varsa, bunlar da aynen aktarılır ve alıcı hücre bu genlerin taşıdığı özel karakterler yönünden pozitif hale gelebilirler. Ancak, bazı durumlarda da, yabancı DNA sekansları, alıcının baz sıralarını değiştirerek veya genleri bölerek mutasyonlara yol açabilir.
Sitoplazmaya ulaşıp ta alıcının kromozomu ile birleşmeyen sekanslar, alıcının endonukleaseları tarafından kolayca parçalanabilir ve etkinliği böylece giderilmiş olur.
Yabancı DNA sekanslarının alıcının kromozomu ile integre olabilmesi başlıca 3 mekanizma ile gerçekleşmektedir.
1) Homolog (genel) rekombinasyon
2) Site specific recombination (özel rekombinasyon)
3) Nonhomolog rekombinasyon
05.01. Homolog (Genel) Rekombinasyon
Bu tarz rekombinasyonda, alıcının hücre duvarından ve belirli yerlerinden içeri giren lineer formdaki DNA segmentleri hemen sirküler forma dönüştürülür. Böyle bir molekülde bulunan ve alıcının kromozomu ile komplementer (homolog) olan bölgeler karşı karşıya gelirler. Homolog bölgelerde meydana gelen karşılıklı kopmalar sonunda, vericinin sekansları alıcının DNA sekansları arasında krosoverla (crossover) integre olur.
Homolog rekombinasyonda rec A+ gen ürünü proteinlerinin önemi oldukça fazladır. Bu gende oluşan mutasyonlar veya bu geni taşımayan rec A- mutantlarında homolog rekombinasyon meydana gelememektedir. Bu genin, aynı zamanda bakterinin UV-ışınlarına ve X-ışınlarına duyarlı hale gelmesinde de rolü vardır. Genel rekombinasyonda, rec A'nın yanı sıra, rec B ve rec C gen ürünlerinin de etkinliği bulunmaktadır. Bu son iki genin inaktivasyonu, rekombinasyon oranını %1 kadar azaltabileceği bildirilmiştir. Bunların dışında diğer bazı genlerin de rekombinasyonda görevi olduğu belirtilmektedir.
Yandaki şekilde rekombinasyon türleri gösterilmektedir. | 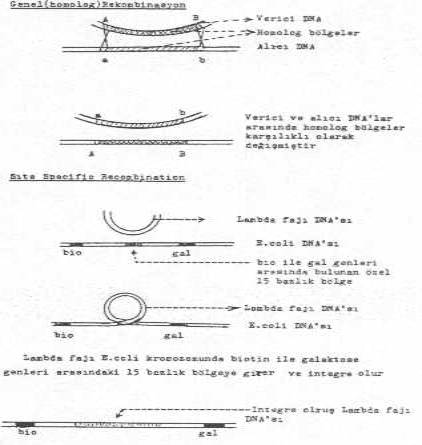
|
Homolog rekombinasyonun yukarda açıklandığı gibi çift iplikçikli iki DNA sekansı arasında (alıcı-verici) olabileceği gibi (Holliday modeli), tek iplikçik ekzogenotun da rekombinasyon yapabileceği belirtilmiştir. Şöyle ki, hücre duvarından tek iplikçik DNA sekansı halinde sitoplazmaya girdikten sonra alıcının homolog bölgesi ile birleşmeden önce çift iplikçikli sirküler hale getirilir ve sonra birleşir veya tek iplikçik şeklinde alıcının homolog bölgesine girer ve burada çift iplikçikli forma dönüştürülür.
Genel (homolog) rekombinasyonda, ekzogenottaki homolog bölge ile alıcının DNA'sındaki homolog bölgeler karşılıklı yer değiştirir. Bazen de alıcının homolog bölgesi sitoplazmaya geçer ve burada endonukleaseler tarafından ayrıştırılır. Dışarıda (sitoplazmada) kalan ekzogenotun homolog olmayan ve integrasyona dahil olmayan bölgeleri de aynı şekilde enzimlerle parçalanır.
Mikroorganizmalar arasında başlıca genel rekombinasyon türleri aşağıda çok kısa olarak bildirilmiştir.
1) Bakteri X bakteri rekombinasyonu: Bakteriler arasında konjugasyon, transformasyon ve transdüksiyon suretiyle aktarılan ve bazı spesifik karakterler taşıyan DNA segmentleri (genler) alıcı hücreye girdikten sonra, hücre DNA'sı ile integre olduğuna gösteren bir çok çalışmalar bulunmaktadır.
2) Faj X faj rekombinasyonu: Bu tarz fajlar arası genel rekombinasyon E. coli 'nin lambda, T2, T4 ve i Æ174 fajlarında rastlanmıştır.
a) Lambda X Lambda rekombinasyonu: E. coli 'yi aynı anda infekte eden iki farklı özellik taşıyan lambda faj DNA'larının homolog bölgeleri arasında kopmalar ve karşılıklı birleşmeler (cross over) sonu rekombinant faj DNA'sı meydana gelir.
b) T2 X T2 rekombinasyonu: E. coli 'ye ait bazı karakterler yönünden birbirinden farklı olan T2 fajları (h- r+ ve h+ r-) aynı anda E. coli 'yi infekte ettiklerinde, katı ortamda rekombinasyon gösteren 4 tür farklı plaklar meydana gelmektedir.(h-r+, h+r+, h-r-, h+r+)
c) T4 X T4 rekombinasyonu: E. coli 'ye ait T4 fajları, arasında da genel rekombinasyonun varlığı gösterilmiştir. Biri 32p ve diğeri de 15N 13C ile işaretli iki T4 fajı ile infekte edilmiş E. coli 'de her iki karaktere sahip fajlar elde edilebilmiştir.
d) Ø X 174 X Ø X 174 rekombinasyonları: Tek iplikçikli sirküler olan bu faj türünde de benzer rekombinasyona rastlanmıştır. Bu fajların hücre içindeki sirküler formları arasında genel rekombinasyonun varlığı saptanmıştır.
3) Plasmid X plasmid rekombinasyonu: E. coli 'ye ait olan Col E1 plasmidleri, hücre içinde genel rekombinasyona maruz kaldıkları gösterilmiştir. Önce, plasmidler homolog noktada 8 rakamı şeklinde birleşirler. Sonra, birbirine temas eden karşılıklı iki halkada kopma meydana gelerek rekombinasyon gerçekleşir.
Yukarıda bahsedilen ve hücre içinde meydana gelen plasmid-plasmid rekombinasyonu, hücre dışında yani in vitro olarak da gerçekleştirilebilir. Rekombinant DNA teknolojisi yardımıyla iki plasmid birleştirilerek yeni rekombinant plasmid meydana getirilebilir ve bu, bakteri içine transfer edebilir.
Plasmidlerle transpozonlar birlikte bulunabilir.
4) Bakteri X plasmid rekombinasyonları: E. coli 'ye ait olan F-faktörü (fertilite faktörü), bu bakteri DNA'sı üzerinde 10'dan fazla homolog bölgeye integre olabildiği ve bakteriyi Hfr (high frequency recombination) haline getirdiği saptanmıştır.
5) Virus X virus rekombinasyonu: Lineer DNA'ya sahip viruslarda (adenovirus, herpesvirus, poxvirus), bunların temperatür sensitif (ts) mutantları veya ts plak tipi mutantları arasında genetik rekombinasyonlara rastlanılmıştır. Rekombinasyon, aynı zamanda, aynı grup içinde olup ta farklı tipteki viruslar arasında da (Adenovirus-tip-2 ile tip-5) görülmüştür.
RNA karakterinde genetik materyale sahip viruslarda (Influenzae virusu) rekombinasyon gözlemlenmiştir. Özellikle, segmentli genoma sahip olanlarda %50'ye varan bir sıklıkta bu fenomen görülmektedir. Segmentler arasında oluşan rekombinasyonlar, segment reassortmenlerine yol açmakta ve bunun sonucunda da antijenik varyasyonlar meydana gelmektedir.
05.02. Site Specific (Özel ) Recombination
Bu tür rekombinasyon, recA gen ürünü proteinine bağımlı değildir. Bu nedenle de recA- mutantlarında da rekombinasyon meydana gelmektedir. Bakterilerin kromozomları üzerinde, bazı yabancı genler için özel alıcı bölgeler bulunmakta ve buradan integrasyon gerçekleşmektedir. Bu tür rekombinasyona en iyi örnek E. coli K-12'ye ait Lambda (l) fajı verilebilir. Bu faj lizojenik formda, bakteri kromozomunda biotin ile galaktoz genleri arasında bulunan 15 bp'lik özel bölgeye integre olur. Faj DNA'sının bu bölgeye girmesi için fajda bulunan int gen ürünü proteinine (integrase) ihtiyaç vardır. İntegrase DNA'da bu özel bölgede çentik açabilir ve kapatabilir.
E. coli K-12 kromozomu üzerinde biotin ile galaktoz genleri arasındaki bu spesifik bölge att (attachment lambda) ile lambda fajındaki attB bölgelerinde oluşan kopmalar, faj DNA'sının kolayca girişini sağlar. Bu girişi integrase enzimi katalize eder. Çapraz bağlanma ile giren faj DNA'sı, kromozomun bir parçası ve devamı haline gelir (profaj). Birleşen yerler DNA ligase ile kapatılır. Bu integrasyon aynı zamanda lizojeniyi de oluşturur.
05.03. Nonhomolog Rekombinasyon
DNA segmentleri arasında genetik homolojiye dayanmadan birleşmeler nonhomolog rekombinasyonlar olarak tanımlanmaktadır. Bu tür rekombinasyonlar, recA gen ürünü proteinlerine de bağımlı değildirler. Bakterilerde rastlanan Transpozonlar (Tn) ve İnsersiyon sekansları (İS-elementleri), bakteri veya plasmid üzerinde veya karşılıklı yer değiştirmeleri bu tür rekombinasyon için örnek kabul edilmektedir.(Mutator faj).
Bakteriyofaj Mu faj (mutator faj, 38 kb) da aynı tarzda yer değiştirebilen genetik materyaller arasında bulunmaktadır.
Bu tür rekombinasyonun meydana geliş mekanizması hakkında özlü bilgiler "Transpozonlar ve İS-elementleri" bahsinde verilmiştir.
06. Prionlar ve Replikasyonları
İnsanlarda (Kuru, CJD, GSS, FFI) ve hayvanlarda (Scrapie, BSE, TME, FSE, vd) Transmissible Spongiform Ensefalopati (TSE)'lere Prion proteini (PrP) olarak tanımlanan ajanların neden olduğu açıklanmıştır. Bu etkenlerin 20 nm'den küçük oldukları ve 254 amino asitlik sekanslara sahip oldukları belirtilmiştir.
Ajan, DNA veya RNA gibi, bakteri veya viruslarda bulunan genetik materyallerin hiç birine sahip olmaması, yüksek ısıya dayanması, DNase ve RNase'lardan etkilenmemesi nedeniyle virus veya bakteri olarak kabul edilmemektedir. Etkenin proteinaz K, iyonizan ışınlara, UV-ışınlarına, yüksek ısıya (134-138°C ve 60 dk), nukleazlara (DNase, RNase) ve bazı dezenfektanlara (formol, Beta-propiolakton) dirençli, buna karşın proteolitik enzimlere duyarlı olduğu açıklanmıştır.
Elektron mikroskopla yapılan çalışmalarda, BSE'li hayvanların beyninde, aynen Scrapie'li koyunlarınkinde rastlanan SAF'ların (Scrapie associated fibrin) eş değeri olan amiloid fibril (amiloid plaklar, amiloid proteinler, amiloid flament) kümeleri oluşur (10-20 x 100-250 nm boyutlarında). Bu fibrillerin hastalık oluşturma yetenekleri vardır ve bunlar prion proteinlerinin (PrPSC) de esasını oluştururlar. Bu amiloid flamentlerden elde edilen 33-37 kDa'luk major proteinlerin aminoasitlerini kodlayan DNA baz sıraları (prion protein geni) saptanmıştır. DNA analizlerinden hareket ederek hazırlanan işaretli (32P) oligonukleotid problar yardımı ile (Prob hibridizasyon) genin yeri belirlenmiştir (insanlarda 20. ve farelerde 2. kromozomda), Bu prion proteini geni, koyunlarda Sip geninin (Scrapie incubation period) eş değeridir. Bu gen (Prn geni), hayvanlardaki transmissible ensefalopatilerde inkubasyon süresini kontrol eden proteinleri kodlamaktadır.
Amiloid fibrillerden hazırlanan proteinlere karşı oluşturulan antiserumların SAF'ları presipite ettiği ve bunların infeksiyöz yeteneklerini nötralize ettikleri açıklanmış ve bu antikorların da spongiform ensefalitik hayvanların beyninde lokalize olabilecekleri belirtilmiştir.
Koyun ve keçilerin doğal Scrapie hastalıkları arasında farkın olması, buna karşın BSE'nin doğal ve deneysel infeksiyonlarında bir uniformitenin bulunması, Scrapie ile BSA hastalığı arasında bir suş karakteristiğinin olabileceğini ve hatta, bunlar ile, insanlardaki spongiform ensefalopatilerin etkenleri arasında da bir farkın bulunabileceği üzerinde durulmaktadır. BSE'li hayvanlardan (sığırlardan), sığırlara yapılan deneysel transferler sonunda oluşan hastalığın tablosu, ile, Scrapie'li koyunların infekte materyallerinden yapılan infeksiyon denemelerinde meydana gelen hastalığın klinik belirtileri arasında azda olsa bir değişikliğin bulunduğu açıklanmıştır. Bu durum, BSE ile Scrapie ajanların arasında da bir suş farkının olabileceği görüşünü ortaya koymaktadır.
Hastalığın nedeni üzerinde araştırıcılar bazı görüşler ileri sürmüşlerdir. Bunlar arasında virus, virion, prion proteini kombine ajan ve genetik bozukluklar en önemlilerini oluşturmaktadır. Ancak, bunlardan prion protein görüşü, deneysel olarak da destek gördüğünden, en fazla taraftar toplayan ajan niteliğini taşımaktadır.
Fare embriyolarına mutant Sip geni sekansları verilerek oluşturulan transjenik farelerde spongiform ensefalopatilerin meydana gelmesi bu genin hastalıktaki rolü ve etkinliğini kanıtladığı gibi, konjenital Prn-p geni bulunmayan farelerde hastalığın oluşturulamaması da,yine, bu genin önemini ortaya koymaktadır.
Prion proteinlerinin vücutta replikasyonlarını izah etmede bazı görüşler ileri sürülmüştür. Bunlardan iki tanesi aşağıda özet olarak verilmektedir.
1) İnfeksiyon teorisi: Vücuda giren patojenik prion proteini (PrPSC), normal prion protein geni (PrP geni, Prn-p) ürünü olan PrPC ile birleşerek bir heterodimer molekül (PrPSC-PrPc) meydana getirir. Bu birleşme sonunda, normal sellüler protein de (PrPc), konformasyonal değişiklikler oluşarak patojenik veya abnormal isoform PrPSC haline dönüşür. Böylece, 2 tane PrPSC veya abnormal isoform PrPSC meydana gelir. Bu iki molekül birbirinden ayrılarak her biri tekrar birer PrPc ile bağlanarak bunları patojenik PrPSC'ye çevirir. Böylece, 4 tane PrPc ortaya çıkar. Bunlarda benzer tarzda replikasyona devam ederler. Bu tarz replikasyon sonunda vücutta PrPSC'ler birikirler. Bunların, afinitesi nedeniyle beyinde toplanması toksik etki meydana getirerek semptomların görülmesine neden olurlar.
İnfeksiyon teorisi yandaki şekilde gösterilmektedir. | 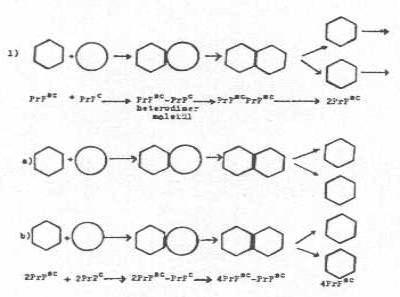
|
2) Genetik teori: Normal hücrelerde prion proteinlerini (PrPc) kodlayan gen, PrP geni (Prn-p geni) bulunmaktadır. Bu gen, insanlarda 20. kromozom ve farelerde de 2. kromozomun kısa kolu üzerinde lokalize olmuştur. Bu gende meydana gelen mutasyonlar (polimorfizm), mutant Prn-p geninin ve dolayısıyla da mutant PrPc'nin oluşmasına yol açarlar. Orijinalinden değişik baz sıralarına sahip olan bu gen, fraklı karakterde amino asitlerin kodlarına sahip olduklarından, protein zincirine değişik amino asitlerin girmesine neden olur. Bu mutant polipeptid, vücutta uzun veya kısa bir zaman süreci içinde, spontan konformasyonal değişikliklere maruz kalarak, patojenik isoform PrPSC'lerin oluşmasına yol açarlar. Bu değişikliğin meydana gelme zamanına göre inkubasyon süresi uzun veya kısa olur.
Genetik teori yandaki şekilde gösterilmektedir. | 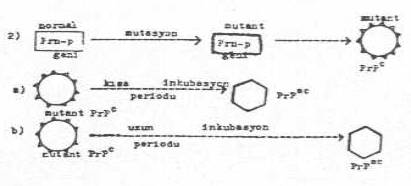
|
1] Kaynak: Temel Mikrobiyoloji