Bakterilerde Varyasyonlar
Prof. Dr. Mustafa Arda
Ankara Üniversitesi Veteriner Fakültesi
Temel Mikrobiyoloji 1
01. Giriş
02. Fenotipik Varyasyonlar (Modifikasyonlar )
02.01. Morfolojik Varyasyonlar
02.02. Kültür Varyasyonları
02.03. Fizyolojik ve Biyokimyasal Varyasyonlar
03. Genotipik Varyasyonlar (Mutasyonlar)
03.01. Genel Bilgiler
03.02. Mutasyonların Başlıca Nedenleri
04. Mutajenik Maddeler (Mutajenler)
04.01. Fiziksel Mutajenler
04.02. Kimyasal Mutajenler
05. Bazı Önemli Mutasyon Türleri
05.01. Spontan (Doğal) Mutasyonlar
05.02. Silent Mutasyon
05.03. Geri Mutasyon ve Reversiyon
05.04. Yapay (Suni) Mutasyonlar
05.05. Mutasyonun Sonuçları
06. Mutant Türleri
06.01. Rezistans Mutantlar
06.02. Nutrisyonel Mutantlar
06.03. Fermentasyon Mutantlar
06.04. Pigmentasyon Mutantları
06.05. İndüksiyon Mutantları
06.06. Antijenik Mutantlar
06.07. Mutasyon Oranı
07. Mutasyonların Tamiri
07.01. Kontrol Okuma ile Düzeltme
07.02. Represör Mutasyonlar
07.03. Dimerizasyonun Giderilmesi
01.Giriş
Mikroorganizmaları identifiye etmede, bunların kültürel, morfolojik, fizyolojik, biyokimyasal ve antijenik özelliklerinden yararlanılmaktadır. Bu karakterler, genellikle, sabit olmasına rağmen, bazı koşullar altında değişmekte ve orijinal mikroorganizmalardan bir veya birkaç yönden ayrı özellikte, yeni türler (varyantlar) meydana gelmektedir. Örn, Salmonellalar, genellikle laktoz negatiftirler. Fakat, kültürlerde 10-6-10-10 oranında meydana gelen değişik örneklerin laktozu fermente ettiğine rastlanabilir. Kapsüllü mikroplar, devamlı pasajları yapılırsa, bu kapsül oluşturma özelliklerini kaybederler. Aynı tarzda, pigmentasyon kabiliyeti de pasajla azalır. Yeni izole edilen (S) karakterindeki mikroplar pasajla R-şekline dönüşebilir ve hastalık yapma kabiliyetlerini kaybedebilirler. Mikroorganizmalarda oluşan bu tür değişikliklere varyasyon adı verilmektedir.
Varyasyonların bir kısmı çevresel koşulların (ısı, ışık, pH, rutubet, osmotik basınç, oksijen azlığı, yüzey gerilimi, antimikrobiyel maddeler, metabolit intermedierler, v.s.) etkisi altında meydana gelirler. Uygun olmayan ve olumsuz yönde etkileyen bu koşullar düzelirse veya düzeltilirse, bakteriler eski formlarına ve karakterlerine dönerler. Genetik düzeyde olmayan ve gelecek kuşaklara aktarılmayan bu tür varyasyonlara modifikasyon (veya fenotipik varyasyon) adı verilir. Modifikasyonlar daha ziyade, kültürel morfolojik ve fizyolojik karakterlerde belirirler.
Bazı değişmeler de, bakteri DNA'sını oluşturan polinukleotid iplikçiklerinde bulunan ve genetik kodları taşıyan nitrojen bazlarının sıralarında meydana gelir. Bu tür değişmeler, genetik düzeyde olduğundan nesillere aktarılır ve devam ederler. Böyle değişmelere de mutasyon (veya genotipik varyasyon) denilir. Mutasyonlar, kendilerini daha çok biyokimyasal, patojenik ve antijenik özelliklerde belli ederler. Böyle değişmeler sonu oluşan ve parental hücrelerden farklı karakter gösteren yeni nesillere mutant adı verilir.
02. Fenotipik Varyasyonlar (Modifikasyonlar )
Fenotipik varyasyonlar, genellikle, optimal çevresel koşulların değişmesi sonu kültürlerde spontan olarak oluşabildiği gibi, normal şartlar altında da meydana gelmektedirler. Besi yerlerinin sınırlı olması nedeni ile kısa bir süre içinde üreyen mikroorganizmalar gıda maddelerini tükettiği gibi ortamda metabolizma artıkları ve toksik intermedierlerin birikmesine, oksijenin sarf edilmesine, osmotik basınç ve yüzey geriliminin değişmesine, kültürlerin eskimesine ve pH'ın düşmesi gibi olumsuz yönde etkileyen koşulların meydana gelmesine neden olurlar. Bu değişen şartlar mikroorganizmalarda 10-6-10-10 oranında modifikasyonlara yol açarlar. Bakterilerde görülen fenotipik varyasyonlar, çeşitli karakterlere etkiler ve orijinalinden farklı nitelikte varyantları meydana getirirler.
Modifikasyonlar, etkilediği başlıca karakterlere göre, şöyle klasifiye edilebilirler.
1- Morfolojik varyasyonlar,
2- Kültür varyasyonları,
3- Fizyolojik ve biyokimyasal varyasyonlar.
02.01. Morfolojik Varyasyonlar
1- Koloni varyasyonları: Eskimiş sıvı kültürlerden, durma veya ölme döneminde olanlarından, katı besi yerlerine ekim yapılırsa başlıca 2 tür koloni karakterine rastlanılır. Bunlardan biri yuvarlak, düzgün, pürüzsüz, parlak ve konveks (S-tipi), diğerleri ise düzensiz, pürüzlü ve mat (R-tipi) kolonilerdir. Bunların dışında ara koloni tipleri (intermedier koloniler ve mukoid-M) koloniler de oluşabilirler.
Eskimiş kültürlerde veya çok pasajı yapılmış mikroorganizmalarda, R-koloni formasyonlarına genellikle, rastlamak mümkündür. Ayrıca, besi yerlerine, Lithium chloride veya anti-S immunserum katılması S ® R varyasyonlarına neden olabilir. Halbuki, R-formundaki mikropları, S-formuna döndürmek daha güçtür. Bunu sağlamak, yani S-formuna döndürmek için, duyarlı laboratuar hayvanlarında pasaj yapılması gereklidir. Mikroorganizmalar S-formundan R-formuna döndükleri zaman birçok karakterinde de değişmeler meydana gelir. Bu durum aşağıdaki tabloda belirtilmiştir.

Mikroorganizmalar, penisilin ve diğer kimyasal ajanların bulunduğu ortamlarda üretilirse, katı besi yerlerinde, normalinden çok değişik, ortası papillalı koloniler oluşur (L-formlu koloni). Bu mikroplar üzerinden baskı kaldırılırsa tekrar normal koloni ve individüel (bireysel) formalarına ulaşırlar.
2- Kapsül varyasyonları: Bazı mikroorganizmalarda (B. anthracis, streptokok, P. multocida, D. pneumoniae, K. pneumoniae, C. welchii, v.s.), hücre duvarının dışında ve bundan ayrı olarak bakteriyi çevreleyen, kalınlıkları ve yapıları türlere göre değişen kapsül formasyonuna rastlanır. Kapsül oluşumu, genellikle, vücut içinde meydana gelmesine karşın, bazı özel koşullar (serumlu, sütlü, karbondioksitli, v.s.) altında vücut dışında da teşekkül edebilmektedir. Kapsül, mikropların antijenik kabiliyetini oluşturduğu gibi virulensi artırıcı özelliğe de sahiptir. Ayrıca, bakteriyi fagositozdan ve diğer bakterisidal vücut maddelerinden de koruma görevi vardır. Mukoid veya S-karakterli koloniler oluşturan kapsüllü mikroorganizmalar, laboratuarlarda uzun süre pasaj yapılırsa veya içinde kapsüler antiserum bulunan besi yerlerinde üretilirse bu yeteneğini yitirirler ve kapsülsüz formlara dönerler. Bu değişiklik ile birlikte hastalık yapma özelliğini de kaybederler. Böyle mikroplar hayvan pasajları ile tekrar kapsüllenirler ve hastalık oluşturma kabiliyetlerini de kazanırlar.
3- Flagella varyasyonları: Mikroorganizmalarda flagella oluşumundaki değişmelere sıkça rastlanılmakta ve flagellalı mikroplardan flagellasız varyantlar meydana gelmektedir. Örn, P. vulgaris ve E. coli 'de bu tür varyasyonlara fazla tesadüf edilebilir. Salmonellalar, içinde %0.1 oranında fenol bulunan ortamlarda üretilirse, flagella sentezi geriler ve flagellasız örnekler meydana gelebilir. Bunlar, normal besi yerlerine aktarılırsa hemen flagellalı formlarını alırlar. L. monocytogenes oda ısısında (22-25°C) üretilirse, 37°C. 'de üretilen kültürlerden daha fazla aktif harekete rastlanır. Çünkü, oda ısısında üreyen kültürlerde, etkende 4 flagella oluşmasına karşın 37°C'dekilerde ise bir tane flagella sentezlenir. Flagellanın kaybı ile, bakteriler H-antijenik özelliklerini ve hareket kabiliyetlerini de yitirirler.
4- Fimbria varyasyonları: Flagellalı veya flagellasız mikroorganizmalarda görülebilen, kısa ve düz fimbrialar (pilus) bu mikropların anaerobik koşullarda, katı besi yerlerinde veya çalkalama kültürlerinin yapılması hallerinde, sentezleri durur ve fimbriasız türler meydana gelir.
5- Spor varyasyonları: Bazı mikroorganizmaların (B. anthracis, B. cereus, B. subtilis, C. tetani, C. welchii, C. botulinum, v.s) in vitro veya in vivo spor oluşturma kabiliyetleri vardır. Sporulasyon her ne kadar bir genetik karakter ise de, oluşumunda çevresel koşulların etkisi de çok büyüktür. Örn, B. anthracis vücut içinde spor vermez, in vitro koşullarda sporulasyona rastlanır. Buna karşın, klostridum sınıfı mikroorganizmalar, anaerobik koşullarda hem vücut içinde ve hem de vücut dışında spor verebilirler. Besi yerlerinde gıdaların azalması sporulasyonu hızlandırır.
6- Şekil varyasyonları: Taze kültürlerdeki veya üreme dönemindeki mikroorganizmalar morfolojik yönden bir örneklilik gösterdikleri gibi, diğer fizyolojik ve biyokimyasal karakterler bakımından da az çok homojen bir durumdadırlar. Kültürlerin eskimesi, bileşiminin değişmesi ve diğer optimal çevresel faktörlerin normallerinden ayrılması sonu, böyle ortamda bulunan mikroorganizmaların şekillerinde bozukluklar (involusyon formları) meydana gelir. Bu formlar kendini, şekillerinin yuvarlak, oval, granüllü, yıldız, halka, flamentli, branşlı, v.s. olmasıyla belli ederler.
Bazı mikroplar da (PPLO), normal üreme dönemi sırasında düzensiz formlar (yuvarlak, halka, yüzük, yıldız v.s.) gösterirler (pleomorfizm).
Penisilin veya anti bakteriyel maddelerin etkisi altında bazı bakterilerin (Streptobacillus moniliformis, v.s.) gerek bireysel ve gerekse koloni morfolojilerinde değişmeler meydana gelebilir (L-formları). Bu mikroorganizmalar kimyasal maddelerin baskısından kurtarılırsa, eski normal form ve kültürel karakterlerini kazanırlar. L-formları, koloni ve bireysel morfolojileri bakımından PPLO 'lara benzerler.
02.02. Kültür Varyasyonları
Mikroorganizmaların sıvı ve katı besi yerlerinde üreme özellikleri ortam karakterinin optimalden ayrılması sonu değişebilir. Örn, B. subtilis, sıvı ortamda genellikle üstte pelikül oluşturarak ürer. Bu ortamın yüzey gerilimi düşürülürse, bu sefer homojen bir tarzda üreme gösterir. S. aureus, sıvı besi yerinde homojen ürer, ortamın yüzey gerilimi artırılırsa, üstte üremeye başlar. Mikropların üreme tarzı üzerine besi yerinin bileşimi ve çevresel koşulların etkisi büyüktür.
02.03. Fizyolojik ve Biyokimyasal Varyasyonlar
1- Boyanma özelliğinde varyasyonlar: Taze kültürlerdeki mikroorganizmalar boyanma özelliklerinde bir örneklik (homojenite) göstermesine karşın, eski kültürlerde bu karakterlerinde sapmalar görülmektedir. Örn, Klostridium sınıfı mikroorganizmalar taze kültürlerde kuvvetli Gram pozitif olmasına karşın, eski kültürlerde ise Gram negatifliğe doğru bir eğilim vardır.
2- Pigment varyasyonları: Pigment oluşturan mikroplar, laboratuarda uzun süre pasajları yapılırsa veya uygun olmayan koşullarda üretilirse, pigmentasyonun zayıfladığı ve kaybolduğu görülür: Örn, S. marcescens en iyi aerobik koşullarda ve oda ısısında tipik kırmızı pigment oluşturur. Aynı mikroorganizma 37°C 'de ve anaerobik şartlarda pigment oluşturmaz. S. aureus da sütlü ortamda iyi pigment yapar ve pasajlar bu özelliği azaltır.
3- Granül oluşumunda varyasyonlar: Bazı mikroorganizmalarda besi yerinin bileşimine göre içlerinde lipid, karbonhidrat (nişasta), fosfat granüllerine rastlanır. Bunların azlığı veya çokluğu, besi yerlerindeki lipid, karbonhidrat ve fosfat bileşiklerine bağlıdır.
4. Enzimatik varyasyonlar: Mikroplar, ortamdaki çeşitli türdeki gıda maddelerinden yararlanabilmesi için, bunlara etkileyen değişik özellikteki enzimleri sentez ederler. Bunların bir kısmı devamlı olarak sentezlenirler (yapısal enzimler) ve bu sentezlenme durumu besi yerinin bileşimi ile fazla ilgili değildirler. Buna karşın, bazı enzimler de besi yerinde özel maddelerin varlığına ve bunların stimulasyonuna bağlıdırlar. Örn; E. coli 'deki beta galaktosidase ve galaktosid permease enzimleri böyledirler. Bu enzimler, ortamda laktoz varsa, bunun uyarımı ile sentezlenirler. Laktoz ortamda bitince, enzim sentezi de durur. Böyle enzimlere, indüklenebilen enzimler adı verilir. Bu nedenle, E. coli 'nin glukozu ayrıştırması, laktozu fermente etmesinden daha çabuktur. Çünkü, glukozu ayrıştıran enzimler her zaman sentezlendiği için hazırdırlar ve zaman geçmeden etkileyebilirler. Buna karşın, laktozun fermantasyonu için, buna etkileyen enzimin (beta galaktosidase) sentezi için en azından 30 dakikalık bir süreye gereksinim duyulur.
5. Diğer değişmeler: Mikroorganizmaların fizyolojik karakterlerinde (toksin, toksik substanslar, enzimler, vb. sentezi), diğer özelliklerinde olduğu gibi, aynı şekilde, varyasyonlar meydana gelebilir. Bunların bir kısmı mutasyonlar sonunda da oluşabilirler.
6. Attenüasyon: Mikroorganizmalar, normal koşulların dışında üretildikleri zaman oluşan değişikliklerden yararlanılarak aşılar meydana getirilmektedir. Örn, B. anthracis 42-43°C 'de devamlı pasajı yapılırsa, yalnız kapsül formasyonunu kaybetmez, aynı zamanda, duyarlı hayvanlar için hastalık oluşturma kabiliyetinde de zayıflama görülür (Pasteur'ün aşısı). Aynı şekilde, sığır tüberküloz mikropları, gliserinli safralı patatesli besi yerinde yıllarca pasajı yapıldıktan sonra, insanlar için çok önemli olan ve korunmada büyük yararlar sağlayan tüberküloz aşısı (BCG) haline getirilmiştir.
At vebası virusunu fare beynine adapte edilmek suretiyle, fareler için patojenik, fakat atlar için patojenik olmayan ancak atlarda bağışıklık oluşturan, attenue virus haline dönüştürülmüştür. Pasteur, kuduzlu tavşanın omuriliğini desikatörde (KOH'lı) kurutmak suretiyle virulensini azaltmış ve aşı olarak kullanmıştır. Doku kültürleri, deneme hayvanları embriyolu yumurtalar ve değişik bileşime sahip sıvı ve katı besi yerleri mikroorganizmaların virulensini azaltmak veya değiştirmek amacı ile kullanılmaktadırlar.
03. Genotipik Varyasyonlar (Mutasyonlar)
03.01. Genel Bilgiler
Mutasyonlar, genellikle polinukleotid iplikçiklerinin (DNA) yapısını oluşturan nukleotidlerdeki bazların, bakteri türüne veya cinsine özgü olan, dizilişlerindeki değişiklikler veya bu bazlarda meydana gelen kimyasal bozukluklar, kopmalar, zedelenmeler, v.s. sonu oluşurlar. Aynı tür içinde bulunan mikroorganizmaların DNA'larında, nitrojen bazları belli ve sabit bir sıra içinde bulunurlar ve böyle fertlerin DNA'ları birbirinin homologudurlar. Bu nedenle, aynı tür içinde bulunan mikroorganizmaların çeşitli karakterleri (morfolojik, fizyolojik, kültürel, biyokimyasal, antijenik vs) birbirine benzerler.
Baz dizilişlerine etkileyen bir faktör veya bazlardaki kimyasal bağlantıların karakterlerindeki değişmeler (tautomerik değişmeler) kendini transkripsiyon ve translasyonda belli ederler. Bunun sonucu olarak da mutasyonlar ve değişik karakterlerde yeni nesiller (mutantlar) meydana gelirler.
Bazı mutasyonlar, bir (veya birkaç) nukleotid çiftini kapsar ve bunlar arasında kalabilir. Bazen de, DNA baz sıraları arasından sadece bir baz çıkabilir veya girebilir (nokta mutasyonu). Bu tür mutasyonlar sonu sentezlenen proteinin aktivitesinde önemli bir değişiklik görülmez ve protein normal görevini yapabilir. Nokta mutasyonları bakteriler tarafından kolayca tamir edilebilirler. Ancak, bir geni (genellikle, 1500 nukleotid çifti) içine alan veya daha büyük oranda meydana gelen mutasyonlar (kromozomal mutasyonlar), polipeptid zincirine çok sayıda yanlış amino asitlerin girmesine neden olacağından, sentezlenen proteinin aktivitesi ya hiç olmaz veya değişik yönde olur. Böyle mutasyonların düzeltilmesi imkânsızdır ve genellikle ölümle sona erer (letal mutasyon). Bakterilerde hareketi sağlayan flagella oluşumunu yöneten genlerde meydana gelen bozukluk, bu organelin sentezlenmesine mani olur. Böylece bakteri, flagellasız ve bunun sonucu olarak da hareketsiz hale dönüşür. Ancak, bu mutasyon ölümle son bulmaz. Buna karşın, çok önemli biyokimyasal olayları katazile eden enzimlerin sentezini idare eden genlerde oluşan kromozomal karakterdeki mutasyonlar bakterilerde ölümler oluşturabilirler.
03.02. Mutasyonların Başlıca Nedenleri
Mutasyonların meydana gelmesine neden olan başlıca faktörler şöyle sıralanabilirler:
1- Polinukleotid iplikçiklerindeki normal baz sıraları arasına bir baz çiftinin çıkması (delasyon) veya baz sıraları arasına bir baz çiftinin girmesi (insersiyon) (nokta mutasyonları).
2- Bir baz çiftinin yerini diğer baz çiftinin alması (transisyonel ve transversiyonal mutasyonlar).
3. Aynı polinukleotid iplikçiği üzerinde yan yana bulunan pirimidin bazları arasında özel bağların kurulması (dimerizasyon).
4. Bazlarla şekerler ve şekerlerle fosfatlar arasındaki bağlantıların kopması.
Baz çiftinin çıkması ve girmesi durumu: Bu tarz mutasyonlarda DNA'daki normal baz sıraları arasından bir baz çifti çıkabilir veya baz sıralarına yeni bir baz çifti girebilir.
a)Baz çiftinin çıkması: Bu örnekte, mRNA üzerinde 16. sırada bulunan sitozin (C) çıkmıştır. Buna göre düzenlenen mRNA ve oluşan yanlış sıralı amino asitler gösterilmektedir. Normal durumda polipeptid zinciri serin, arginin, tirosin, cystein, ....le başlamakta ve devam etmektedir. Buna karşın, bir baz çiftinin (sitozin) çıkması sonu oluşan yeni programlanmaya göre, amino asit dizilişi serinden alanin, threonin, alanin tarzında olup sırası olmayan amino asitlerdir. Bu nedenle proteinin karakteri değişik olur veya inaktif bir durum gösterir. Eğer anlamsız kodonlardan biri (UGA, UAG, UAA) sıraya girerse, protein sentezi sona erer. Bazen de, böyle değişmeler aynı amino asite ait diğer 2. veya 3. tripletlerden birini oluşturursa; (Örn; argininin 6 kodonu CGU, CGC, CGA, CGG, AGA, AGG; serinin 4 kodonu ACU, UCC, UCA, UCG veya leucinin 6 kodonu, UUA, UUG, CUU, CUC, CUA, CUG, v.s.) bu sefer polipeptid zincirinde bir süre daha doğru amino asitler sırada bulunabilir. Bu durum, eğer kısa bir süre sonra yeni bir mutasyonla düzeltilirse, proteini inaktif bir molekül olmaktan kurtarır.
DNA’dan bir baz çiftinin çıkması yandaki şekilde gösterilmektedir. | 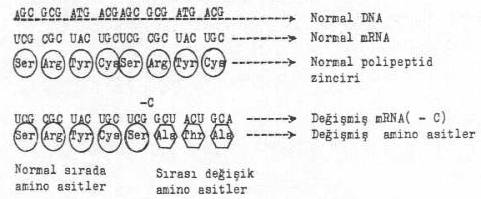
|
b) Baz çiftinin girmesi: Baz çiftinin girmesi de aynı yukarıdaki örnekte olduğu gibi, baz sıralarının değişmesine ve mRNA'nın yeniden programlanmasına yol açacak ve polipeptid zincirinde değişik amino asitlerin araya girmesine neden teşkil edecektir.
DNA’ya bir baz çiftinin girmesi yandaki şekilde gösterilmektedir. | 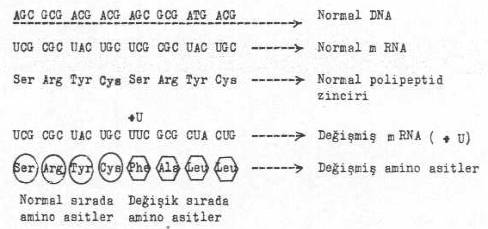
|
Bu örnekte normal mRNA'da 13. sıraya U bazı girmiş ve mRNA yeni baştan programlanmış ve sıraya, Phe + Alanin + leucin + leucin + gibi değişik amino asitler girmiştir. Serinden sonra gelen amino asitler normal polipeptid zincirlerinde sırada değildirler. Bu nedenle oluşan protein inaktif bir durum gösterebilir.
Bir baz çiftinin çıkmasını, aynı gen içinde veya dışında ikinci bir mutasyon aracılığı ile bir baz çiftinin girmesi izleyebilir. Bu durum iki şekilde etkileyebilir, ya birinci mutasyonla oluşan değişmelere, ikinci mutasyonunki de katılır ve öylece tamiri olanaksız mutasyonlar oluşabilir veya ikinci mutasyon, birincinin düzelmesine veya tamirine yol açabilir. Böylece ilk mutasyon giderilir. Bu ikinci mutasyon birinciye ne kadar yakınsa, polipeptid zincirinde sıraya giren yanlış amino asitler de o oranda az olur.
DNA’da baz sıraları arasına birden fazla bazların girmesi ve çıkması durumları yandaki şekilde gösterilmektedir. | 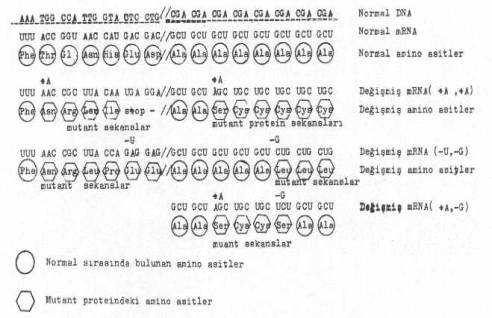
|
Transisyonel mutasyonlar: Bu tür mutasyonlar, genellikle ya spontan olarak veya mutajenik maddelerin etkisiyle oluşurlar. Transisyonel türdeki mutasyonlara, pürin ve primidinlerin elektronlarında oluşan tautomerik değişmelerin bazlar arası karşılıklı hidrojen bağlantısının özelliklerini değiştirmesine neden olur. Örn; timin genellikle keto formunda C = O olup, bu durumu ile adeninle karşılıklı çift hidrojen bağı ile birleşir. Ancak, timin, nadiren enol formuna (C—OH) dönüşürse, 3 bağlantı yerinin oluşması nedeniyle, adeninle değil de, guaninle birleşir. Böylece, DNA molekülünde AT çifti yerine GT çifti geçmiş olur. Ancak, bu durum uzun süre devam etmez ve iki-üç generasyon sonra değişerek, guanin bu sefer sitozinle bağ kurar ve AT ® GC transisyonu meydana gelir. Böylece, başlangıçta AT olan molekül, 2-3 generasyon sonra GC çiftine dönüşmüş olur.
Replikasyon sırasında pürin yerine diğer bir pürinin, veya pirimidin yerine diğer bir pirimidin bazının girmesine transisyonel mutasyon denir. Örn, adenin yerine guanin (AºG) veya sitozin yerine timinin girmesi (CºT) gibi. Eğer pürin yerine pirimidin (veya tersi) girerse buna transversiyonel mutasyon adı verilir. Örn; adenin yerine sitozin (AºC) veya guanin yerine timinin girmesi (GºT).
Eğer baz çifti değişmeleri iki yönlü ise (AT W GC) bunlar bidireksiyonal mutasyonlar olarak isimlendirilir. Bazı mutajenler (hidroksilamin) tek yönlü transisyonel mutasyonlar oluşturur (GC ® AT).
Adeninin amino formunda iken, imine formuna dönüşmesi, aynı şekilde, karşılıklı bağlantıları değiştirir ve sitozinle bağ kurmasına yol açabilir (A—C).
Dimerizasyon: Mikroorganizmalar, ultraviole ışınlarının etkisine maruz kaldıktan sonra, aynı DNA iplikçiğinde yan yana bulunan pirimidinler (genellikle timinler) fotokimyasal reaksiyonlar sonu birbirleri ile kovalent bağlarla birleşirler (T - T). Işığın dozu artarsa bu sefer sitozinler de birleşebilirler (C—C). Dimerizasyon iki nukleotid arasını kısalttığından, DNA'da çarpıklıklara yol açar ve DNA'nın duplikasyon mekanizması bozulur. Sonunda transkripsiyon ve translasyon düzeninde değişiklikler meydana gelir.
DNA'da diğer bozukluklar: Birçok mutajenler DNA iplikçiklerinin çeşitli yerlerinde bozukluklar oluşturabilirler. Bu tür bozukluklar kısaca şöyledir:
a) Baz-şeker bağlarının kopması: İyonizen ışınlar, alkilleyen etkenler, nitroz asiti, hidroksil amin, süksinil peroksit, vs.
b) Bazlar arası hidrojen bağların kopması: Isı, asit, alkali, iyonizan ışınlar, guanidinum klorid, vs.
c) Çapraz bağ teşkili: Alkilleyen etkenler, nitröz asiti, vs.
d) Şeker-fosfat bağlarının kopması: İyonizan ışınlar, sonik ve ultrasonik vibrasyonlar, asit, alkali, suksinil peroksit, vs.
04. Mutajenik Maddeler (Mutajenler)
Bakterilerde mutasyon oluşturan etkenler, genellikle, fiziksel, kimyasal ve biyolojik karakterdedirler. Bunlar da,
04.01. Fiziksel Mutajenler
Isı: Eğer bakteriler 100°C 'ye kadar yavaş yavaş ısıtılırsa iplikçikler arasındaki karşılıklı hidrojen bağları çözülür ve iki iplikçik birbirinden ayrılır (denatürasyon). Isı yavaş yavaş azaltılırsa iki iplikçik hemen birleşir. Bu olay, hibridizasyonda işe yarar. Bilinen baz sıralarına sahip test DNA'sı ile bilinmeyen DNA (veya RNA) iplikçiğinin homologluk durumu araştırılır. Bu durum bakterinin klasifikasyonunda yararlı olur.
Ultraviolet ışınları: Ultraviolet ışınlarına maruz bırakılan bakteriler, primidinler (timin veya sitozin), pürinlerden daha fazla UV ışınlarını absorbe eder ve kendilerinde birçok fotokimyasal değişmeler meydana gelir. Bunun sonucu, atomlar arası enerjinin artmasına ve timin dimerlerinin oluşmasına sebep olur. Bu bozukluk iki baz arası 0,34 nm'lik uzaklığı 0.28 nm.'e indirerek DNA'da çarpıklıklar meydana getirir. Dimerlerin bulunduğu yerlerde transkripsiyonda atlamalar veya boşluklar oluşur ve bu bölge atlanarak devam edilir.
İks (x) ışınları: Dalga boyu UV-ışınlarından çok daha kısa olan X-ışınlarının bazların atomlarında oluşturduğu yüksek enerji, UV-ışınlarınkinden 4 kat daha fazladır. Bazlar tarafından çok çabuk absorbe edilen bu ışınlar yüksek enerjilerinden dolayı, DNA iplikçiklerinden nukleotidlerin çıkmasına ve mutasyonlara sebep olmaktadır. Bu mutasyonlar genellikle ölümle son bulur. İyonizan ışınlar, bakterilerde bazı kimyasal değişiklikler de meydana getirirler. Bunun başında iyonizasyon gelmektedir. İyonizasyon sonu atomların orbital elektronlarında dışarı fırlamalar oluşur. Serbest kalan elektronlar, hücredeki bazı radikaller veya moleküller tarafından alınarak, yeni bileşikler teşekkül eder. Bunların çoğu bakteriye zarar verir niteliktedir.
Ultrasonik vibrasyonlar: Ses ötesi vibrasyonlar da bakteriler üzerine olumsuz yönde etkiler ve bazı genetik değişmeler meydana getirebilirler.
04.02. Kimyasal Mutajenler
Kimyasal mutajenik maddelerin etkisi altında oluşan mutasyonlar kolayca tamir edilebilirler. Mutajenik maddeler, genellikle, transisyonel ve transversiyonel mutasyonlara yol açarlar: Bazıları da bazların çıkmalarına ve DNA'da kopmalara sebep olurlar. Başlıca mutajenik maddeler:
Nitröz asiti (HNO2):
Bazlar üzerine direkt etki yapan nitröz asiti, banlardan oksidatif deaminasyon ile amin grubunu (NH2) çıkarır ve bunun yerine keto (= O) grubunu koyar. Sitozinin de aminasyonu sonu urasil (U), ve adeninin deaminasyonunda da hipoksantin meydana gelir. Bu olaylar sonunda oluşan urasil adeninle ve hipoksantinde sitozinle bağ kurar.
HNO2
Sitozin +¾¾¾® Urasil (urasil — adenin) GC ® AT
HNO2
Adenin +¾¾¾® Hipoksantin (hipoksantin-sitozin) AT ® GC
Nitröz asiti aracılığı ile oluşan mutasyonlar iki yönlüdürler (bidireksiyonal, AT ® GC ® AT).
Hidroksil amin (NH2OH): Bu mutajen daha ziyade pirimidinler (sitozin) üzerine etkiler ve olay sonu oluşan yeni bileşikler, tautomerik değişmelere maruz kalır ve transisyonel mutasyonlar oluşur. Sitozinin tautomerizasyonu sonu adeninle birleşme meydana gelir. Bu transisyonel mutasyon (CG ® AT) tek yönlüdür. Sitozinin, adeninle birleşmesi başlıca 2 şekilde izah edilmektedir. Biri, hidroksil amin, sitozinin 6 pozisyonunda bulunan amin grubu ile reaksiyon vererek bunu Oxime (= N-OH) haline çevirir ve sonra adeninle birleşme olur. Diğeri ise, sitozinin 4,5 durumundaki çift bağla reaksiyon verir ve oluşan bileşik, tautomerik değişmelere maruz kalarak adeninle birleşir.
Alkilan maddeler: Birçok kimyasal maddeler, nukleotidler arasına alkil gruplar (CH3-, CH2-) sokarak transisyonel mutasyonlar yaparlar. Bu tür alkilan maddeler arasında sülfür, nitrojen mustard, dimetil sulfonate (DMS), dietil sulfonate (DES), etil metane sulfonate (EMS), metil metane sulfonate (MMS), nitrosoguanidine (NG), v.s. sayılabilir. Bunlardan, EMS ve NG, timin ve guaninin 6 pozisyonundaki keto grubunu alkile ederek bunları tautomerizasyona iter. Diğer alkilanlar ise, bazların diğer pozisyonlarına etkiler. Değişmiş olan bazları DNA polimerase tanıyamaz ve bu bazlar transkripte olamazlar. Sülfür ve nitrojen mustardlar bundan başka, DNA ipliklerinde çapraz bağlantılara neden olabilir ve ayrıca helikste kopmalar yapabilir.
Baz analogları: Baz analogları parental DNA'ya etkilemezler. Bunlar replikasyon sırasında, yeni iplikçik sentezlenirken, sıraya girecek bazların yerine geçerler (veya bunların yerlerini alırlar) ve böylece transisyonel mutasyonlara yol açarlar. Baz analogları arasında en fazla üzerinde çalışılanı timinin analogu 5- Bromouracil (5-BU) ve adeninin analoğu 2-Amino pürin (2-AP)'dir. Bu analoglar, kimyasal yapı bakımından, bazlara benzediğinden, replikasyon sırasında bazların yerini alabilirler. Timinin analoğu olan 5-BU, keto formunda iken adeninler ve enol formunda iken de guaninle bağ kurar. Sonra guanin de sitozinle bağ kurarak, AT çifti GC çiftine dönüşür (AT ® GC). Yeni oluşan GC baz çifti sonraki generasyonlarda tautomerik değişmeler sonu AT 'ye çevrilirler (GC ® AT).
A=T®A= BU (keto) ® G= BU (enol) G= C (transisyonel çift)
A= BU
G=C®G=BU (enol ® A = BU (keto) ® A=T (transisyonel çift)
Adeninin analoğu olan 2-AP, replikasyon sırasında adeninin yerini alır. Sonradan timinle ve diğer replikasyonlar da tautomerik değişmeler sonu sitozinle bağ kurarak transisyona yol açar. Sitosin de, sonradan guaninle birleşir.
A= T ® T = AP ® C º AP ® C º G
Böylece AT çifti CG 'ye dönüşür. Diğer baz analoglarından, timidin analoğu 5- Bromodeoxy uridine, A-T bağlantısı olan yerlere replikasyon sırasında etkilidir. Aynı şekilde 5- Fluoro deoxyuridine tymidylate senthetase enzimi üzerine inhibitör etki yapar.
Akridinler: Akridin boyaları arasında en iyi bilineni proflavindir. Proflavin tarafından oluşturulan mutasyonlar, baz analogları, nitröz asiti ve hidroksilamin tarafından geri çevrilemez. Ancak, mutasyon olan gende spontan mutasyonlar veya tekrar akridin boyaları ile oluşturulan ikinci bir mutasyon ile düzeltilebilir. Proflavin, DNA baz çiftleri arasına girerek en azından 1-20 adet baz çiftinin çıkmasına sebep olabileceği gibi, bazı durumlarda, baz çiftinin girmesine de neden olabilir. Akridin boyaları, bakterilerde olduğu gibi, fajlarda da mutasyonlar oluşturabilir.
Diğer mutajenler: Metilkolantren, MnCl, arsenik, krom, urethane,creosol, katran, organik peroksidase, asitler, alkaliler. İndirekt etkileyen mutajenler: İlaçlar, hormonlar, fazla oksijen, pH değişmeleri, diğer çevresel faktörler.
Biyolojik Mutajenler: Bakterilerde bulunan bazı ekstra kromozomal genetik elementler (plasmid, faj, transpozon, Mu fajı, İs-elementleri) mutasyonlara yol açabilirler.
Bunlar hakkında ileriki bahislerde gerekli bilgiler verilmektedir.
05. Bazı Önemli Mutasyon Türleri
05.01. Spontan (Doğal) Mutasyonlar
Bakteriyel populasyon, dışarıdan herhangi bir indükleyici madde katılmadan veya müdahale edilmeden, mutasyonlara maruz kalabilir. Buna spontan mutasyon adı verilmektedir. Oluşan mutantlara da spontan mutantlar denilir. Bakteri DNA'ları çoğalma sırasında çok fazla replikasyona tabi tutulmaktadır. Doğaldır ki, çok sayıda ve sınırsız replikasyonlar ve DNA'nın diğer fonksiyonları sırasında, nukleotidlerin asamblesinde ve polimerizasyonunda bazı hatalar meydana gelmektedir. Bu hatalara, bir bazdaki elektronların tautomerik transpozisyonu oldukça fazla neden olmaktadır. Örn; timin normal olarak keto durumunda bulunur ve adeninle karşılıklı iki hidrojen bağı kurar. Eğer, timin, DNA'nın replikasyonu sırasında, enol forma değişirse, bu zaman guaninle birleşir. Bu durumda, yeni DNA'da AT yerine GC bazları girmiş olur.
Timinin keto ve enol formları yandaki şekilde gösterilmektedir. | 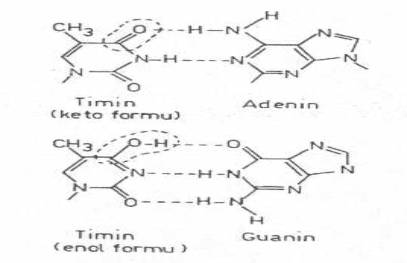
|
05.02. Silent Mutasyon
Moleküler düzeyde, DNA'da bazlarda meydana gelen her türlü değişiklik mutasyon olarak kabul edilirse de, böyle mutasyonlar her zaman fenotipik olarak eksprese edilmezler. Bazı tripletlerin 3. bazında meydana gelen değişmeler fenotipe etkilemez. Bakteride hiç bir aksaklık meydana getirmez. Böyle değişmelere silent mutasyon adı verilir. Bazen de bir tripletin 2. veya 1. bazları da değişse bakteride önemli bir aksaklığa yol açmayabilir. AUC 'nin AUG'ye dönüşmesi isoleucin yerine met'in girmesine neden olur. Ancak, böyle bir mutasyon bir sorun yaratmadığı halde eğer, CUU yerine CCU girerse, leucine yerine prolinin girmesine yol açar ki, böyle bir değişiklik polipeptid zincirinde konformasyon üzerine önemli derecede etkiler.
05.03. Geri Mutasyon ve Reversiyon
Mutantlar, kendi DNA'larında oluşan bazı mutasyonlar sonu tekrar orijinal (parental) veya doğal formlarını kazanabilirler (geri mutasyon). Bu olguda, değişen amino asit tripleti, geri mutasyonla tekrar orijinal amino asiti kodlayan triplet haline gelebilir.
Böylece, bakteriler tekrar orijinal fenotipik karakterlerine kavuşurlar. Böyle durumlara, aynı gende oluşan supresör mutasyonlar (intragenik) veya kromozom üzerinde başka gende meydana gelen mutasyonlar (ekstragenik supresörler) yol açabilirler.
05.04. Yapay (Suni) Mutasyonlar
Eğer hücreler mutajenik ajanlarla (kimyasal, fiziksel, biyolojik, vs.). muamele edilirse, mutantlar meydana gelebilir. Bu tarzda başlıca 4 tür mutant oluşabilmektedir.
1- Bir baz çiftinin diğeri ile yer değiştirmesi, örn., AT yerine GC 'nin girmesi gibi,
2- Bir baz çiftinin girmesi veya çıkması,
3- Büyük bir DNA segmentinin (bir veya birkaç genlik) kaybolması veya kromozom üzerinde başka bölgeye transferi (transpozisyonlar).
4- Biyoteknolojik yöntemlerle mutasyonlar oluşturulabilir ve mutant suşlar elde edilebilir.
05.05. Mutasyonun Sonuçları
Mutasyonların mikroorganizmalar üzerinde ne denli etkilediği başlıca iki faktör tarafından belirlenir. Bunlardan biri, gen ürünlerinin ne kadar değiştiği ve diğeri de gen ürünlerinin hücre içi ne derece de önemli olduğudur.
Gen ürünlerindeki değişiklikler: Bütün mutasyonlar, genetik kodları ve bunlara bağlı olarak da genetik bilgileri (informasyonları) değiştirir. Ancak, bunların gen ürünleri üzerindeki etkileri farklıdır. Bazıları hücre fenotipinde çok az veya hiç bir değişiklik yapmaz. Örn; bir kodonda meydana gelen değişiklik sonu oluşan gen ürünü, normal kodonla aynı olabilir. Bir amino asit bir kaç kodonla temsil edildiği için, böyle bir mutasyonun hiç bir zararlı etkisi yoktur. Bazen de değişiklik sonu oluşan farklı amino asitin gen ürününün fonksiyonuna bir zarar vermez. Farklı bir amino asitin şifresini oluşturan mutasyonlara genellikle, "missense mutasyon" adı verilir.
Bazen, böyle olgular hücrelerde büyük değişikler veya ölümler meydana getirebilir. Gen tümden inaktive olabilir veya gen ürünü fonksiyonel olmayabilir.
Bazen de mutasyonlar, hiç bir amino asit kodlamayan kodonlar UAG UAA UGA meydana getirebilir. Örn; stop kodonu gibi. Bunlara da "nonsense mutasyon" adı verilir.
Esansiyal gen ürünleri: Bazı gen ürünleri hücrenin yaşamı için çok önemlidir. Bunların olmamasının hücrede oluşturduğu bozukluklar, gen ürünün fonksiyonlarına bağlıdır. Örn; DNA veya RNA polimerase enzimini kodlayan gende meydana gelen bozukluklar, bu enzimlerin fonksiyonlarını bozduğundan hücrenin yaşamına da etkiler. Hücreler çoğalamazlar.
06. Mutant Türleri
Gerek spontan mutasyonlar (bilinen mutajenlerin olmadığı durumlarda, uygun olmayan çevresel koşulların altında kendiliğinden oluşan ve daha ziyade girme, çıkma, transisyon ve transversiyon tarzında görülen mutasyonlar) ve gerekse fiziksel veya kimyasal mutajenlerin etkisi altında oluşan mutasyonlar sonu, birçok karakterlerde değişmeler meydana gelebilir. Böyle mutasyonlar sonu oluşan başlıca mutant türleri şöyledir.
06.01. Rezistans Mutantlar
Mutasyonlar sonu çeşitli ilaç, dezenfektan, antibiyotik, kemoterapötik, inhibitörler, ultraviole ışınları, fajlar, v.s. etkenlere karşı dirençlilik gösteren mutantlar oluşabilir.
06.02. Nutrisyonel Mutantlar
Özel üretme faktörlerine gereksinim göstermeyen orijinal parental (prototrof) hücrelerden, mutasyonlar sonu bir veya birkaç üretme faktörüne gereksinim duyan mutat (okzotrof) hücreler meydana gelebilir. Prototrof hücreler basit ortamlarda (minimal ortam) gelişebildikleri halde okzotroflar gelişemezler. Bunların besi yerlerine üretme faktörleri (amino asit, vitamin, nukleik asit, v.s.) katılır (kompleks ortam).
06.03. Fermentasyon Mutantlar
Orijinlerinde karbonhidratlardan bazılarını fermente etmeyen (veya eden) suşlardan oluşan mutantların bu karbonhidratlara etkilediği (veya etkilemediği) görülebilir. Örn; salmonella, proteus veya shigella arasında laktozu fermente eden, E. coli arasında da laktozu fermente etmeyen mutantlar ortaya çıkabilir.
06.04. Pigmentasyon Mutantları
Bazı mutasyonlar, pigment oluşturan mikroorganizmalarda bu kabiliyetin kaybolmasına yol açar. S. marcescens veya S. aureus 'dan uzun pasajlar veya mutasyonlar sonunda pigment oluşturmayan mutantlar elde edilmiştir.
06.05. İndüksiyon Mutantları
Bu tür mutantlar, mutajenik maddelerin etkisiyle meydana gelirler.
06.06. Antijenik Mutantlar
Bakterilerde bulunan kapsül, flagella, hücre duvarı ve piluslar antijenik karakterlere sahiptirler. Bunların biyosentezini idare eden genlerde oluşan mutasyonlar, bu faktörlerin meydana gelmesine ve fonksiyonlarına etkiler ve değişikliğe uğratarak antijenik bozukluklara yol açarlar. Flagellalı bakteriler flagellasız, ve kapsüllüler de kapsülsüz hale gelebilirler. Somatik (O) antijenlerinde de önemli değişmeler görülür. Hücre duvarında lokalize olan bu antijen salmonellalarda lipopolisakkarid fraksiyonda lokalize olmuştur. Bu lippolisakkaridin iç kısmında 2-keto 3-deoxyoctanate, etanolamin, fosfat, lipid ve dış kısmında da kısa zincirli glikoz, galaktoz, heptozfosfat ve N-acetyl glukoz amin yerleşmiştir. Spesifik yan zincirlerde bulunan şekerlerin cinsi ve tekrarlanan ünitelerdeki dizilişleri, salmonellalar arasında O-antijenik özelliği yaratır. Esas yapıda veya yan zincirlerin oluşumunu idare eden genlerde meydana gelen mutasyonlar; O-antijenik özelliğinin değişmesine veya kaybolmasına neden olur. Böyle mutasyonlar sonu S-kolonilerinden R-tipli koloniler meydana gelir. Ayrıca, yan zincirlerin tekrarlanan üniteleri polimerize olmasa, yine mutantlar ortaya çıkabilir.
Hücre duvarının biyosentezlerine çeşitli dönemlerde etkileyen mutasyonlar, canlılığa tesir etmeden, antijenik karakterde değişmeler yapabilir.
Bazen flagella hareketini yöneten gende mutasyon olabilir. Böyle bakterilerde flagella olsa bile hareket olmaz. S. typhimurium 'da pürin sentezine etkileyen mutasyonlar, bu mikrobun fareler için virulensinin kaybolmasına neden olurlar.
06.07. Mutasyon Oranı
Kültürlerde oluşan spontan mutasyonlar az çok sabit bir durum gösterebilir. Mutasyon oranı her generasyonda, her bir hücreye isabet eden mutasyon miktarı ile ölçülür. Mutasyonlar genellikle, replikasyon sırasında ve yeni sentezlenen iplikçiklerde meydana gelirler. Parental DNA'da mutasyon çok azdır.
Mutasyon oranı (MO) aşağıdaki şekilde hesaplanır.
Ms
MO = ¾¾¾¾¾
N1-No
MO: Mutasyon oranı
Ms: Mutasyon sayısı
N0: Başlangıçtaki mikrop sayısı
N1: Bir generasyon sonra mikrop sayısı
Yukarıdaki formül yanı sıra, mutasyon sıklığını hesaplamada Poisson dağılımı formülünden de fazlaca yararlanılır.
P(x)= ( mx/X!) e¾m
e: Doğal logaritma (2.178)
m: Her petrideki (tüpteki) ortalama mutant sayısı
Bu formüle göre;
1- Hiç mutant ihtiva etmeyen tüp sayısı m=o),
P(0)= e-m veya P(0) = -m 'dir.
2- Bir mutant ihtiva eden tüp sayısı (m = 1)
P(1) = e-1 veya P(1) = 1/e = 0.37'dir.
07. Mutasyonların Tamiri
Spontan olarak veya mutajenik maddelerin etkisiyle oluşan mutasyonlar, bakterilerde özel mekanizmalar tarafından tamir edilebilir ve düzeltilebilir. Ancak, bu işlem, oluşan bozukluğun büyüklüğüne ve önemine göre değişebilir. Nokta mutasyonları genellikle ve kolayca giderilebilirse de, bir genin kaybolması ve kromozomda oluşan kopmalar kolayca tamir edilemez ve ölümle son bulurlar.
Mutasyonların tamir mekanizması, DNA'da oluşan bozuklukların karakterine göre değişir. Bu da birkaç tarzda görülebilir:
07.01. Kontrol Okuma ile Düzeltme
DNA'nın sentezi (replikasyon) sırasında yanlış nukleotidlerin sıraya girmesi hücre tarafından hemen fark edilerek, ileri sentez çok kısa bir süre için durdurulur. DNA polimerase I ve III'ün geriye doğru kontrol okuma mekanizması ile hatanın yeri bulunarak yanlış nukleotid 3-OH ucundan hidrolize edilerek çıkarılır ve yerine doğru nukleotidler konarak bozukluk giderilir.
07.02. Represör Mutasyonlar
Eğer DNA'da baz çiftleri arasına bir bazın girmesi (veya çıkması) ile oluşan mutasyon, aynı genin içinde (intragenik) veya gen dışında (ekstragenik) meydana gelen ve diğer bir baz çiftinin çıkması (veya girmesi) şeklinde oluşan ikinci bir mutasyonla, giderilebilir (supresör mutasyon). İkinci mutasyon, birinciye ne kadar yakınsa, düzeltme işlemi hem kolay ve hem de imkân dahilinde olur. Ayrı genlerde oluşan ikinci mutasyon, birinci ile arasında çok mesafe olacağından ve bu arayı yanlış sıralı aminoasitler dolduracağından, tamiri olanaksızdır ve bakterinin ölümüne sebep olur.
Supresör mutasyon, birinci mutasyonun etkisiyle bozulan aminoasit sıralarını düzeltir, polipeptid zincirine normal sıralı aminoasitlerin girmesine ve proteinin aktivite kazanmasına yol açar. Supresör mutasyon da başlıca üç tarzda çalışabilir. 1- Birinci mutasyon sonu değişen baz sıraları, aynı yerde oluşan ikinci bir mutasyonla düzeltilebilir. Örn, ilk mutasyonda GC baz çiftinin yerine AT baz çifti girmişse aynı yerde oluşan ikinci mutasyon, son giren baz çiftinin çıkmasına yol açar ve ilk mutasyonu ve etkisini ortadan kaldırır. 2- İlk mutasyonla DNA'ya giren baz çifti, ikinci bir mutasyonla, bağlanma sırası değiştirilir ve hata düzeltilir. Örn, ilk mutasyon GC yerine AT 'yi koymuşsa, ikinci mutasyon AT bağlantısını TA şekline çevirerek oluşan bozukluğu, tam olmamakla beraber kısmen giderebilir. Bu şekildeki tamir sonu oluşan mutant orijinalinden farklıdır. Ancak, enzim aktivitesinde pek önemli değişiklik görülmeyebilir. 3- İkinci mutasyonun aynı gen içinde veya gen dışında meydana gelmesi ile ilk bozukluk düzeltilebilir.
İntragenik ve ekstragenik mutasyonlar genellikle, farklı veya yanlış sıralı amino asitleri (sırada olmaması gereken) kodlayan, yeni kodonları devreye soktuğu gibi (missense mutasyon), bazı hallerde bir amino asitin karşılığı olmayan kodonlar (UGA, UAA, UGA) oluşturmakta, bunlar da mRNA üzerinde sıraya girmektedirler (anlamsız mutasyon, nonsense mutasyon). Birincide, normal sıraya girmesi gereken aminoasit yerine, oluşan mutasyon sonu değişen baz sırası nedeniyle, başka bir aminoasit kodonu gelir ve bunlar değişik aminoasitlerin polipeptid zincirinde sıraya girmesine neden olur ve proteinin aktivitesini bozar. Buna karşılık, anlamsız mutasyonda ise, değişen baz sırası nedeniyle meydana gelen yeni kodon, herhangi bir amino asiti kodlamaz veya bunun aminoasit alfabesinde karşılığı olan aminoasit bulunmaz. Böyle 3 tane bulunan kodonlardan biri UGA, UAG, UAA protein sentezi sırasında mRNA üzerinde sıraya girerse, polipeptid sentezi ve uzanması durur.
İntragenik mutasyonların, nokta mutasyonları tarzında ve sadece bir nukleotid çiftinde oluşması, tamiratını kolaylaştırmaktadır. Çünkü, birinci mutasyon ile ikinci mutasyon yeri arasında fazla bir mesafe yoktur. Bozukluğun tamiri ikinci mutasyonun oluşma süresine bağlıdır. Bu süre uzun olursa, yanlış (farklı) aminoasitler fazla miktarda polipeptid zincirine girebilir. Eğer bir generasyon sonra oluşursa, yeni nesilde herhangi bir bozukluk oluşmaz. Çünkü, tüm aminoasit sıraları düzelmiştir. Örn, bir gende alanin kodonu (GCC) bulunsun ve primer mutasyon sonu bu kodon değişerek valin (GUC) haline getirilsin. Bu ilk mutasyon sonu oluşan ve yeni mutantlarda sentezlenen protein, inaktif bir durumda olabilir. Bundan sonra oluşan ikinci bir mutasyon, alanini aktive eden enzimin yapısını tayin eden gende meydana gelirse, bu son mutasyon, enzimin, tekrar alanini tanıyarak onu aktive etmesini ve kendine özel alanin tRNA'ya bağlanmasını sağlar ve böylece ilk mutasyon düzeltilir veya aksine, eğer primer mutasyon alaninin yerine valini sıraya koymuşsa, ikinci supresör mutasyon valin tRNA'nın yapısını tayin eden gende oluşursa, bu sefer, alanini aktive eden enzim, tRNA'yı alaninle yükler hale getirebilir ve mutasyonu düzeltilebilir.
Sentetik mesenger polyuridylic asit polipeptid zincirine fenil alanini girmesine neden olur. Çünkü, bunda UUU kodonu vardır. Streptomycinin bulunduğu ortamda ise, mesenger polyuridylic acid, isoleusin (AUU), serin (UCU) ve diğer aminoasitlerin polipeptide girmesine yol açar. Steptomycinin (veya buna kimyaca yakın antibiyotiklerin) bu etkisi, bu antibiyotiğin ribosomlara bağlanması ve kodonun okunmasını değiştirmesi ile oluşur.
P. mirabilis normal olarak streptomycine duyarlıdır (Ss). Bundan oluşan bazı mutantlar, ancak, streptomycin bulunan ortamlarda üreyebilirler. Diğer bir deyimle, üremeleri streptymocine bağlıdır (Sd). Bu tarz oluşan varyasyon kısa bir süre sonra değişir ve mutantlar normal orijinal fonksiyonel durumlarına dönerler (Ss).
07.03. Dimerizasyonun Giderilmesi
Ultraviolet ışınlarının DNA'da bulunan primidinler tarafından absorbe olması ve enerji ile yüklenmesi sonu oluşan dimerizasyon (polinukleotid iplikçiğinde yan yana bulunan pirimidinlerin birbirleriyle birleşmesi) primidinlerin karşı iplikçikteki pürinlerle bağlanmasını önler ve DNA'yı çarpıtır. Bu durum replikasyona ve bunun sonucu olarak ta transkripsiyona ve translasyona etkileyerek mutasyonlara yol açar. Dimerizasyon başlıca 3 tarzda giderilir:
1- Fotoreaktivasyon: Eğer bakteriler, UV-ışınlamasından sonra, hemen görülebilen ışınlara tutulursa, UV-ışınlarının letal etkisi giderilir. Bu olayda UV-ışınlaması sırasında oluşan timin dimerleri (T-T), gün ışığında aktive olan ve iş görebilen özel enzimler (fotoreaktif enzimler) tarafından hidrolize edilerek aralarındaki bağlantı koparılır. Böylece timinlerin, normal ve karşı iplikçikteki pürinlerle bağ kurması temin edilir.
2- Karanlıkta reaktivasyon: Bu tarz tamirat, ışıkta tamirattan farklıdır. Bu mekanizma ışıkta çalışmaz ve başlıca 4 enzim (endonuklease, ekzonuklease, polimerase, ligase) sıra ile iş görerek düzeltme görevini yaparlar. Düzelme olayı şu basamakları izler.
Karanlıkta reaktivasyon ve aşamaları yandaki şekilde gösterilmektedir. | 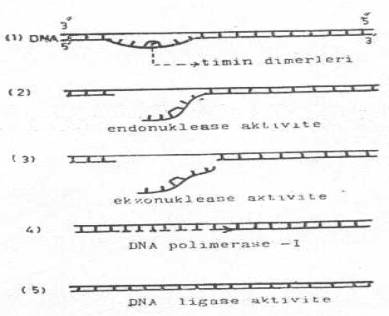
|
a) Dimer bulunan bölge endonuklease enzimleri tarafından sınırlandırılır ve ayrılır (koparılır).
b) Ekzonuklease enzimleri tarafından kenarları biraz daha genişletilir ve dimerler sindirilir.
c) DNA polimerase-I, sağlam karşı iplikçiği örnek (kalıp) olarak kullanılarak, bozuk veya kopuk yerde 5'®3' yönünde hemen ikinci komplementer iplikçiği sentezler.
d) Oluşan yeni iplikçiğin uçlar, polinukleotid ligease enzimi yardımı ile eski iplikçiğin uçlarına birleştirilir ve böylece tamirat yapılmış ve dimerler giderilmiş olur.
Dimerlerin enzimatik yolla giderilmesine bazı kimyasal maddeler (caffein gibi) mani olabilir. UV-ışınları etkisine maruz kalan faj, böyle bir tamirat mekanizmasına sahip bakteriye girince, reaktive olur ve eski durumunu kazanır. Eğer, UV-ışınlarına maruz kalan faj, irradiye edilmiş diğer bir bakteriye girerse, çok sayıda fajın reaktive olmasına ve dışarı çıkmasına neden olur (UV-reaktivasyon).
3) Endonukleotik insizyonla tamir: Hücrelerde (prokaryotik ve ökaryotik) bulunan bazı endonukleaselar de DNA'daki lezyonları tanıyarak bunlardaki fosfodiester bağlarını kaparak çıkarılmasını katalize ederler. Bu fonksiyona sahip E. coli 'de insizyon endonukleazların (I, II, III;,IV, V, VI) varlığı açıklanmıştır.
4) Rekombinasyonla tamir: DNA'da meydana gelen bozuklukların giderilmesinde rekombinasyonla tamir mekanizması da görev yapmaktadır. E. coli 'de UV-ışınlarının oluşturduğu dimerlerin tamirinde bu sistemin de iş gördüğü açıklanmıştır. Replikasyonda atlanan dimer bölgesi, karşı parental DNA segmenti tarafından rekombinasyonal tamir mekanizması ile kapatılır sonra DNA'daki timin dimerleri de eksizyonla çıkarılır. Bunların yerleri de DNA polimerase ve DNA ligase tarafından düzeltilir.
5) Bypass ile tamir: Bu mekanizma, lezyonu tam olarak ortadan kaldıramaz ve lezyon atlanarak replikasyona devam edilir. Eğer, bozukluk fazla ise hücreler ölebilir. Eğer sıraya giren yeni baz tamiratı düzeltirse, bozukluk ta fazla ileri gitmeden ortadan kaldırılmış olur.
6) N-glikosidase ile tamir: Bu mekanizmada, DNA'da bulunan anormal bazlar (urasil, hipoksantin, 3-metiladenin, v.s.) N-glikosidase enziminin katalitik etkisiyle N-glikosilik bağları koparılarak çıkarılır. Bir çok türde N-glikosidase enzimi belirlenmiştir. Örn, N-3, N-7 metil ve etil purinleri kaldıran spesifik enzimler gibi. Bazı mikroorganizmalar (E. coli, B. subtilis, M. luteus, vs) ve memeli hücrelerinde de (dana timusu ve insan hücreleri) bu tarz aktivite bulunduğu açıklanmıştır. Sitozinin deamine olması sonu meydana gelen urasil, N-glikosidase enziminin aktivitesi sonu çıkarılarak hata düzeltilir. M. luteus 'da timin dimerlerinin tamirinde N-glikosidase enziminin aktivitesinin önemli fonksiyonu olduğu belirlenmiştir. Bu mekanizmada enzim, dimerlerin N-glikosilik bağlantısını koparır.
7) Dealkilasyonla tamir: Bazı mikroorganizmalarda, mitojenik veya karsinojenik etkiye sahip olan bazı maddelerin (alkilan maddeler vs) etkisiyle kimyasal olarak modifiye olan (alkillenen) bazları tanıyarak bunları dealkile etmek (veya çıkartmak) suretiyle hatayı düzelten enzim sistemlerinin varlığı saptanmıştır. Alkilan bir madde olan dimetilnitrozamin kuvvetli karsinojenler arasında bulunmaktadır. Farelere verildiğinde, DNA'nın çeşitli yerlerinde metilasyonlar meydana getiren bazı bileşikler oluşturur. Metile olan DNA ürünü (O6 - metilguanin), böbrek ve karaciğerde bulunan spesifik bazı enzimler tarafından kaldırılarak hata tamir edilir.
1] Kaynak: Temel Mikrobiyoloji