Klonlamada Kullanılan Başlıca Vektörler
Prof. Dr. Mustafa Arda
Ankara Üniversitesi Veteriner Fakültesi
Temel Mikrobiyoloji 1
01. Genel Bilgiler
02. Plasmid Vektörler
02.01. Normal Plasmid Vektörler
02.02. Ekspresyon ve Ekskresyon Vektörler
02.03. Mekik (shuttle) Plasmid Vektörler
02.04. Plasmid Tabanlı Ökaryotik Vektörler
03. Faj Vektörler
03.01. Lambda Fajı
03.02. M13 Fajı
03.03. Kosmid (cosmid) Vektörler
04. Viral Vektörler
04.01. DNA Virus Vektörleri
04.02. RNA Virus Vektörleri
05. Bakteriyel Vektörler
01. Genel Bilgiler
Mikroorganizmalar arasında gen klonlamasında bir çok aracı moleküllerden (vektör) yararlanılmaktadır. Bunların amaca yönelik ve bilinçli olarak seçilmesi ve kullanılması gereklidir. Özellikle, restriksiyon haritalarının, baz sıralarının, gen fonksiyonlarının, hücre içi replikasyon mekanizmalarının, spesifik markerlerinin, vs. diğer özelliklerinin çok iyi bilinmesi ve tanınması klonlamanın başarısı için ilk adımı oluşturmaktadır.
Gen klonlamasında başlıca 4 temel vektörden yararlanılmaktadır. Bunlar da,
1) Plasmid vektörler,
2) Faj vektörler,
3) Viral vektörler,
4) Bakteriyel vektörler.
02. Plasmid Vektörler
Bazı prokaryotik (bakteriler) ve ökaryotik organizmalarda (maya, flamentöz mantar, mavi algler, vs.), hücrenin büyük kromozomundan ayrı olarak sitoplasma içinde ya serbest olarak (plasmid) veya hücre kromozomu ile birleşmiş halde (episom), DNA karakterinde ekstrakromozomal genetik elementler bulunmaktadır. Doğal olarak bulunabilen bu tür plasmidlere, E. coli, P. aeruginosa, S. aureus, B. subtilis. B. anthracis, C. perfringens, C. tetani, C. botulinum, vs. sıkça rastlanılmakta ve bunların bazıları da virulensle çok ilişkili bulunmaktadırlar. Mayalardan, S. cerevisiae ’de de böyle plasmidlere (2 µ DNA plasmidi) tesadüf edilmiştir. Doğal olarak bulunan bu plasmidlerden gen aktarımında pek fazla yararlanılamamakta, bunların yerine in vitro koşullarda ve amaca göre hazırlanan suni plasmidler tercih edilmektedirler.
Plasmidler, çok sayıda, kolay ve ucuz elde edilmeleri, çift iplikcikli, sarmal ve sirküler bir yapıya sahip olmaları, yapılarında seleksiyonda önemli fonksiyonu olan bazı spesifik markerlerin (ampr, telr, kanr, lac+, vs.) bulunması, belli uzunlukta yabancı genleri kabul etmeleri, infeksiyöz olmamaları, restriksiyon endonukleaz için bazılarında bir tek kesim yerinin bulunması, replikasyon orijinine sahip olmaları ve buna bağlı olarak ta hücre içinde bağımsız olarak replikasyon özelliği taşıması, vs. gibi hususlar plasmidlerin fazla oranda vektör olarak kullanılmalarının başlıca nedenleri arasında gelmektedir.
Plasmidlerin DNA’larında bulunan böyle bölgeler (tet, ampr, kanr, strr, lac+, vs.), bunların hücre içinde replikasyonları için pek önemli değildirler. İşte, rekombinant plasmid (kimerik plasmid, hibrid plasmid) elde etmede, bu sekanslar içine aktarılması istenen gen (hedef gen, hedef DNA) yerleştirilir ve hangi genin içine transfer edilmişse o gen inaktif hale gelir ve diğer gen(ler) aktivitelerini korurlar.
E. coli ‘ye ait plazmidde değişik RE’ler için kesim yerleri yandaki şekilde gösterilmektedir. | 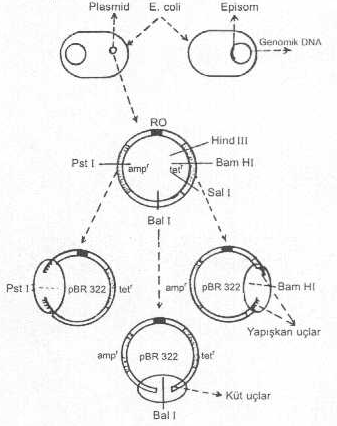
|
Rekombinant DNA teknolojisinde kullanılan suni plasmid vektörler yapılarına ve fonksiyonlarına göre başlıca 4 temel grupta toplanabilirler. Bunlar da;
02.01. Normal Plasmid Vektörler
Bu tür plasmid vektörlerde sadece rezistenslik veya diğer genler (ampr, tetr, lac+, vs.) ile replikasyon orijinleri vardır. Basit bir restriksiyon haritasına sahiptirler. Yabancı gen DNA segmenti, rezistenslik genlerinden biri içine aktarılarak inaktive edilir ve diğer gen, koloni seleksiyonu için, sağlam olarak bırakılır. Plasmidle birleştirilen gen, sonra, alıcı hücrelere (Örn., E. coli ’ye) aktarılır. Alıcı hücrede genin eksprese edilip edilmediği, gen ürününün kontrolü ile yapılır. Ancak, bu tarzda hazırlanan vektörlerdeki genler alıcı hücre içinde çok zayıf eksprese olurlar veya olmayabilirler. Bu amaçla, pBR322 plasmidinden yararlanılabilir.
02.02. Ekspresyon ve Ekskresyon Vektörler
Yabancı gen DNA’sının alıcı hücrede (Örn., E. coli ’de) etkin bir tarzda ekspresyonunu sağlamak için bakterinin replikasyon mekanizması tarafından kolayca tanınabilen ve alıcı bakteriye ait kuvvetli bir prokaryotik promotorlardan (DNA’nın üzerinde bulunan ve RNA polimeraz enziminin bağlandığı ve transkripsiyonun başladığı bölge) biri yabancı gen DNA’sının 5'- ucuna bağlanarak plasmide eklenir ve böyle hazırlanan vektör plasmid, alıcı hücre tarafından kolayca algılanarak ekspresyon şansı, normal plasmid vektörlere oranla, çok artmış bir duruma getirilir. En fazla kullanılan promotorlar arasında, tryptophan promotoru, beta galactosidase vs. sayılabilir. Bu tarzda oluşturulan plasmidlere ekspresyon plasmidleri adı verilir.
Bazı durumlarda da promotorun ön kısmına, diğer bazı DNA sekansları (Shine-Dalgarno sekansları) ilave edilerek, transkripsiyonda oluşan mRNA’ nın ribosomun 30S alt ünitesine güvenle ve kuvvetle başlanması garanti altına alınır. Böyle plasmidlere ayrıca AUG başlatma kodonu ile terminasyon sekansları da bağlanabilir. Bu amaçla UGA tripleti kullanılır.
Bir ekspresyon plazmidi yandaki şekilde gösterilmektedir. | 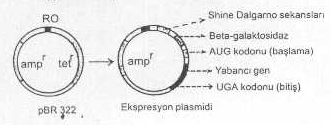
|
Alıcı bakteri içinde, ekspresyonunun sağlaması istenen gen ürününün makromolekül olduğu (Örn., IgG Fc-porsiyonu, insülin, vs.) durumlarda, sentezlenen substanslar hücreden dışarı çıkamaz ve içerde kalır. Bunun kısmen de olsa hücre dışına salgılanmasını sağlayabilmek için ekskresyon plasmid vektörler geliştirilmiştir. Örn., insan IgG’nin Fc-porsiyonunu kodlayan geni, Bacillus sp. ait beta laktamaz geni ile birlikte bir plasmide (pEXFC-10) bağlandıktan sonra E. coli ’ye transfer edilirse, E. coli içinde beta laktamaz enzimi sentezlenip dışarı salgılanırken IgG’nin Fc-porsiyonu da dışarı çıkar. Aslında, bu plasmidler de, bir tür ekspresyon plasmidleridir .
Bir ekspresyon vektörü yandaki şekilde gösterilmektedir. | 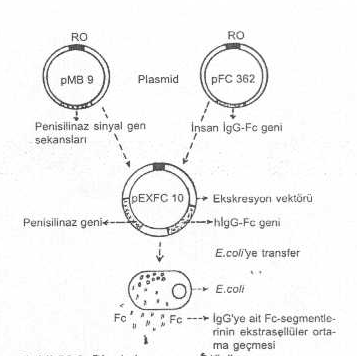
|
02.03. Mekik (shuttle) Plasmid Vektörler
Yukarıdaki tarzda dizaynı yapılarak kullanılan plasmid vektörler (normal veya ekspresyon vektörleri) sadece bir tür alıcı hücrede veya konakcıda replike olabilir ve taşıdıkları gen eksprese edilebilir. Bunun dışındaki hücrelerde (ökaryotiklerde) replike olamazlar. Çünkü, bu son alıcı hücrelere ait RO'ya ve promotora sahip değildirler. Eğer, prokaryotik promotor taşıyan ekspresyon vektörlerine, ökaryotiklere ait bir promotor da ilave edilirse, bu tarzda hazırlanan vektör hem prokaryotik ve hem de ökaryotik mikroorganizmalarda replike olabilir. Örn., prokaryotik promotor içeren plasmide, Saccharomyces cerevisiae ’ye ait bir promotor olan Leu-2 (veya tryptophan, histidin, arginin, vs.) de ilave edilirse, bu plasmid hem bakteride ve hem de maya hücresinde replike olabilir ve taşıdığı genin ekspresyonunu sağlayabilir (bifonksiyonel plasmidler).
Bir mekik plazmidinin genel yapısı yandaki şekilde gösterilmektedir. | 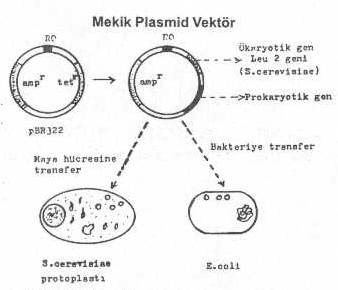
|
Gen konstraktlarının az miktarda hazırlandığı veya gen DNA’sının az elde edildiği durumlarda, birinci alıcı hücrede (Örn. E. coli ’de) genin ekspresyonu değil de, genin fazlaca üretimi ve gen sayısının artırılması (amplifikasyon) amaçlanabilir. O zaman, kendisine yabancı gen taşıyan plasmid aktarılmış olan E. coli, içinde chloramphenicol (170 mg/ml) bulunan ortamda üretilirse, bakteride çok sayıda (binlerce) spesifik gen taşıyan plasmid gelişebilir ve bakteri DNA’sının % 30-40'ına ulaşabilir. Ayrıca, genin ampflikasyonu için, spesifik primerler kullanılarak PCR’dan da yararlanılabilir. Gen sayısı artırıldıktan sonra bakteri lize edilerek içinden plasmid ekstre edilir ve bundan da hedef gen DNA’sı çıkarılır. Eğer istenirse, bu gen DNA’sı bu mikroorganizmaya ait vektör plasmide eklenerek tekrar E. coli ’ye veya maya hücresine ait bir plasmide (pRITI2363) bağlanarak S. cerevisiae protoplastına transfekte edilebilir. Örn., Hepatitis B virusuna karşı aşı hazırlamada bu yöntemden yararlanılmıştır.
Ekspresyon vektörleri yardımı ile E. coli içinde bir çok genin klonlaması ve ekspresyonu sağlanmıştır. Bunlar arasında, insan insülin hormonu, insan büyüme hormonu, rat büyüme hormonu, sığır büyüme hormonu, interferon, difteri toksini, ürokinaz enzimi, pilus antijeni, ovalbumin,HGPRT enzimi, Hepatitis B, vs. vardır.
02.04. Plasmid Tabanlı Ökaryotik Vektörler
Bu tür vektörler, genellikle, ökaryotik hücrelere (memeli, kanatlı hücreleri) yabancı genlerin aktarılması ve ekspresyonu için geliştirilmiş vektörlerdir. Bu nedenle de, yabancı genle birleştirilmiş olan plasmide, kuvvetli viral promotor ve regulatör (terminasyon) sekanslar da ilave edilerek COS hücrelerine transfekte edilirler. Eğer istenirse, önce, yabancı gen taşıyan plasmid bir bakteriye aktarılarak genin fazlaca üretimi sağlanabilir. Ökaryotik hücreye transfekte edilen vektördeki genin ekspresyonu sağlanır.
03. Faj Vektörler
Gen transferlerinde, aracı diğer bir molekül olarak, çeşitli faj DNA’larından da yararlanılmıştır. Bakteriyofajlar, genel anlamda, bakterilerde üreyen ve lizis oluşturan bakteri viruslarıdır. Bir çok bakterinin kendilerine özgü fajları vardır ve bunlar, bakterilerin identifikasyonlarına ve teşhislerine de büyük katkıda bulunurlar.
Fajların bazıları, kendi konakçılarında lizis (virulent fajlar) meydana getirmelerine karşın bir kısmı da bakteri içinde bulunduğu halde, herhangi bir zararlı etkide bulunmazlar (Örn. E. coli ’ye ait lambda fajı, C. diphtheriae ait beta fajı, vs.). Bu tür fajlar bakterinin genomu ile birleşir, onun bir devamı haline gelir, onunla birlikte eş zamanlı olarak replike olur ve her hücre bölünmesi sırasında da, bakteri DNA’sı ile birlikte kardeş hücreye aktarılır (profaj). Lambda fajı ve beta fajı birer profajdırlar.
Vektör olarak, genellikle, E. coli ’ye ait fajlardan (Lambda, M13. vs.) ve ayrıca fajmid ve kosmidlerden de yararlanılır.
03.01. Lambda Fajı
Gen transferi amacı ile lambda fajı (çift iplikcikli, lineer, yaklaşık 50 kbp uzunlukta, 31 x 106 dalton molekül ağırlığında) başarı ile denenmiştir. Faj DNA’sının her iki ucunda (5'-uçlarında) 12 bazlık ve birbirinin komplementeri olan nukleotidler lokalize olmuşlardır (yapışkan uçlar, cohesive ends, cos).
Faj DNA’sı E. coli ’ye girdikten sonra başlıca 2 replikasyon formu gösterir.
1) Litik form: Faj başlığı içinde lineer tarzda bulunan faj DNA’sı, bakteriye girdikten sonra yapışkan uçlar birbirleriyle birleşerek, molekül sirkülarize olur. Bu form, bakteri içinde, bidireksiyonal olarak hızla çoğalır ve faj DNA’sı artar. Bir süre sonra, bu dairesel replikatif formlardan (RF) bazıları, rolling circle replikasyon fazına geçerek birbirlerine yapışkan uçlarla (coslarla) birleşmiş lineer uzun faj DNA’sı meydana gelir. Bu dönemde fonksiyonu olan özel enzimler (paketleme enzimleri, protein A) lineer faj DNA’sını, cos bölgelerinden, 50 kb’lık mesafelerden olmak üzere, keserek oluşan segmentleri faj başlığı içine paketler. Böylece olgun fajlar meydana gelir ve bakteri parçalanarak fajlar dışarı çıkarlar.
2) Lizojenik form: Bakteri içinde bulunan fajlardan bazıları, E. coli kromozomunda galactose ile biotin genleri arasındaki özel bölgeye homolog rekombinasyonla girerek integre olur ve faj, bakteri DNA’sının bir devamı haline gelir (profaj). Bu faj bakteriyi normal koşullar altında lize etmez ve eş zamanlı olarak replike olur ve kardeş hücreye aktarılır.
Lambda fajının orta bölgesi, bakteri içinde litik bir üreme için gerekli değildir. Bu bölge, başka bir gen ile yer değiştirebilir. Yani, fajın ortası çıkarılarak buraya yabancı gen aktarılabilir. Böyle bir değişme de fajda zararlı bir etki yaratmaz. Ancak, yanlardaki sekanslar önemli olup litik üreme için gerekli bilgileri taşırlar. Ayrıca, her biri 15-20 kb uzunlukta olan yanlarda, faj başlığını ve kuyruğunu kodlayan sekanslar da bulunur.
Fajın orta bölgesi restriksiyon endonukleaz enzimi ile iki yerinden kesilerek orta segment çıkarılır, buraya yaklaşık aynı boyda, yabancı gen taşıyan segment (aynı RE ile kesilmiş) fajın orta kısmına yerleştirilir. Sonra, paketleme enzimi yardımı ile faj başlığı içine paketlenir ve buradan da başka bir E. coli ’ye transfer edilir. Böylece, yabancı bir gen faj vektör yardımı ile bakteriye aktarılmış olur.
Lambda fajının yapısı ve klonlama aşamaları yandaki şekilde gösterilmektedir. | 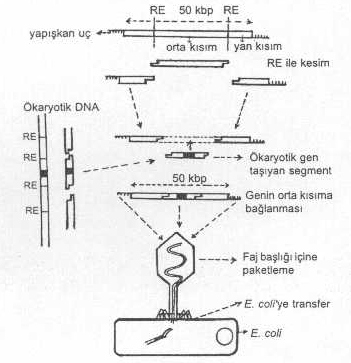
|
Yabancı geni, fajın orta kısmına birleştirmede dikkat edilecek önemli nokta paketleme enziminin tanıma sınırlarıdır. Bu enzim, ancak 50 kb uzunlukta bulunan cos-bölgelerini tanıyarak buralardan keser. Eğer, yabancı gen çok küçük ise, iki cos-bölgesi arası daralacağından, paketleme enziminin tanıma sınırlarının altında kalacaktır veya yabancı gen çok büyükse, aktarılan genle birlikte yanların toplam uzunluğu 50 kb’ın çok üstünde olacağından paketleme enziminin tanıma sınırlarının yine dışında olacak ve böylece, her iki gen DNA’sı fajla birleşse bile faj başlığı içine paketlenemeyecektir. Diğer önemli noktada, gerek faj ve gerekse yabancı gen DNA’sı aynı RE ile kesilmesi ve birbirlerinin komplementeri olan yapışkan uçların meydana getirilmesidir. Yukarıda bahsedilen problemleri gidermek için özel fajlar (lambda gt-beta, lambda gt-10, lambda gt-11, charon faj serisi, vs.) hazırlanmıştır.
03.02. M13 Fajı
Tek iplikcikli flamentöz fajlardan olan E. coli M13 fajı, lambda fajı kadar olmasa bile, yabancı genlerin aktarılmasında denenmiştir. M13 fajı, E. coli ’nin seks pilusuna yapışır ve yaklaşık 6.8 kb uzunluktadır. M13 fajı bakteri içine girince, kendine ait olan tek iplikçik DNA, bakterinin replikasyon mekanizması tarafından buna ikinci bir cDNA sentezlenerek, çift iplikcikli hale dönüştürülür. Böylece sirküler ve çift iplikcikli replikatif formlar (RF) meydana gelir. Bu RF'ler rolling circle tarzında bir replikasyon gösterirler. M13 fajının bir E. coli içinde yaklaşık 100-200 kopyası bulunabilir. Belli bir üreme döneminden sonra, tek iplikcikli faj DNA’sı oluşmaya başlar ve bunlar faj başlığı içine girerler.
M13 fajının replikasyon aşamaları yandaki şekilde gösterilmektedir. | 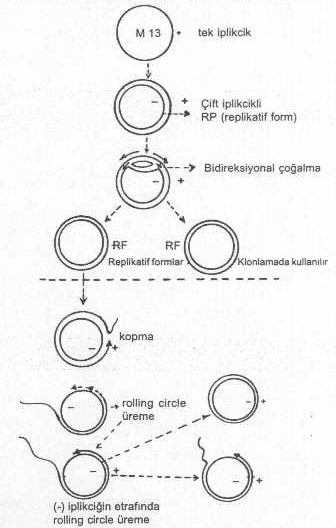
|
M13 fajının çift iplikcikli replikatif formları gen aktarımlarında vektör olarak yararlanılır. Bu formlar, bakteriden ekstre edilerek çıkarılır ve üzerinde gerekli manipulasyonlar yapılır. RF’lere yaklaşık 300-400 bazlık bir yabancı geni aktarmak mümkündür. RF’ler bir restriksiyon endonukleaz ile kesildikten sonra, E. coli ’ye ait beta galactosidase (Z) geni ve bunun da amino terminal bölgesine yakın kısmına polilinker bağlamak mümkündür. Bu işlem, fajda herhangi bir fonksiyonel bozukluk meydana getirmediği gibi, E. coli ’ye transfekte edildiğinde, bu gen eksprese olabilmektedir. Eğer kendisinde aktif beta galactosidase geni bulunan E. coli, bileşiminde, isopropyl-trigalactosidose (IPTG) ve bir kromojenik madde olan X gal bulunan katı ortamda üretilirse, faj bakteriyi eriterek mavi renkli plaklar meydana getirir. Eğer, beta galactosidase geninin bir bölgesinde bulunan polilinker içine bir yabancı gen aktarılırsa, o zaman, bu gen inaktive olur ve bakteri içinde eksprese olamaz ve renksiz plaklar meydana gelir. Bu denemelerde kullanılan E. coli, beta galactosidase yönünden negatif bir karakter taşımaktadır.
Birinci denemede, bakteriye aktarılan beta galactosidase aktif olduğu için sentezlenen enzim ortamdaki substratlarla etkileşerek mavi renk meydana getirmesine karşın, ikinci denemede gen inaktif olduğu ve bakteride de bu gen bulunmadığından renksiz plaklar oluşmaktadır.
03.03. Kosmid (cosmid) Vektörler
Kosmid vektörler, bir bakteriye ait ve replikasyon orijin bölgesine sahip plasmidle, buna birleştirilmiş lambda fajı yapışkan ucundan (cos, cohesive ends) oluşturulmuşlardır. Bazı kosmidlerde tek bir cos bulunmasına karşın, iki bölgeli veya mekik kosmidler meydana getirmek de olanak dahilindedir.
1) Tek cos bölgeli kosmid vektörler: Bu vektörler için lambda fajının E. coli ’de rolling circle tarzındaki replikasyon fazından elde edilen lineer faj DNA’sındaki cos bölgeleri RE’lerle kesilerek çıkarılır. Bunlar, pBR322 plasmidinin tetr geni içine integre edilir ve böylece tek bir cos bölgesine sahip kosmid vektör elde edilir. Böyle bir kosmid yaklaşık 6 kb uzunlukta olup, yapısında plasmide ait replikasyon orijini (RO), ampr geni ve cos bölgesi bulunur. Bu tarzda hazırlanan kosmid yaklaşık 35-40 kb uzunlukta yabancı bir geni kabul edebilir ve aktarabilir. Kosmide yabancı gen integre edebilmek için, önce kosmid Bam HI ile muamele edilir ve molekül açılarak lineer hale getirilir. Alkalin fosfataz ile etkilendirilerek vektör-vektör ligasyon (sirkularizasyon) önlenir. Aynı anda da yabancı gen DNA’sı, aynı RE ile muamele edilerek gen DNA segmenti çıkarılır. Bu parçanın 35-40 kb kadar olması, paketleme enziminin tanıması için gereklidir. Gen DNA’sının iki ucundaki yapışkan uçlarla, açılan bölgedeki yapışkan uçlar birleştirilerek vektör hazırlanır. Paketleme enzimi de 45-50 kb uzaklıktan tanıyarak keser ve faj başlığı içine paketlenir. Bir kosmidde iki tane ampr geni veya iki tane orijin bölgesi bulunamaz.
Kosmid vektör oluşturulması yandaki şekilde gösterilmektedir. | 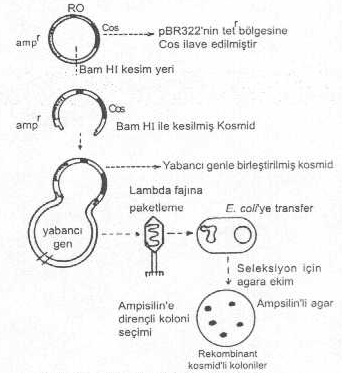
|
2) Çift cos bölgeli kosmid vektörler: Tek cos bölgeli vektörlere bir alternatif olarak iki cos bölgesine ve iki markere (ampr, kanr) sahip vektörler de hazırlanmış ve kullanılmıştır. Kosmidler iki RE ile aynı anda kesildiğinde iki lineer parça elde edilir ve her birinde bir cos bölgesi ve bir marker bulunur. Yabancı gen DNA’sı, iki cos bölgesi arasına integre edilir. diğer işlemler aynen tek cos bölgeli kosmidde olduğu gibi yürütülür.
3) Mekik (shuttle) kosmid vektörler: Bu tarzda dizaynı yapılan vektörlerde cos bölgeleri ve markerleri yanı sıra prokaryotik ve ökaryotik hücrelere ait genetik DNA sekansları da bulunur. Bunlar da 35-40 kb’lık yabancı DNA’yı kabul edebilir ve transferini sağlayabilir.
4) Fajmidler: Plasmid ve fajların bazı sekanslarından oluşan fajmidler de bazı genlerin transferinde denenmiştir. Örn., immunglobulin geninin E. coli ’de ekspresyonunda pHEN10 fajmidinden yararlanılmıştır.
04. Viral Vektörler
Viral vektörler, daha ziyade, prokaryotik ve/veya ökaryotik genlerin hücrelere (memeli, kanatlı, vs.) aktarılması ve genlerin ekspresyonları için, çok uygundurlar. Ökaryotik hücrelerde üredikleri için de böyle vektörlere, ökaryotik viral vektörler adı da verilmektedir. Viral infeksiyonlara karşı etkin aşıların hazırlanmasında da son yıllarda, viral vektörler oldukça önem kazanmıştır.
Virusların vektör olarak kullanılabilmesi için, bunların moleküler yapılarının, hücre ve konakcı spektrumlarının, hücrelerde replikasyon özelliklerinin, paketlenmelerinin, hücrelerde oluşturdukları bozuklukların ve diğer karakterlerinin çok iyi bilinmesi gerekmektedir. Ayrıca, bazı kriterlere de uymaları aranan vasıflar arasındadır. Şöyle ki, konakcı ve hücre spektrumu geniş olmalı, büyük bir DNA segmentini kabul edebilmeli ve kendi replikasyonunu önlememeli, infeksiyöz ve onkojenik olmamalı ve birden fazla farklı genleri alabilmelidir.
Genetik materyallerin ökaryotik hücrelere etkin bir tarzda transferlerini ve ekspresyonlarını sağlamak için bazı DNA ve RNA viruslarından rekombinant ökaryotik viral vektör olarak yararlanılmıştır. Bunlardan başlıcaları aşağıda özet olarak belirtilmiştir.
04.01. DNA Virus Vektörleri
1) Simian virus 40(SV40) vektörleri: DNA karakterinde çift iplikcikli, sarmal ve sirküler bir genoma sahip olan SV40, yaklaşık 5.2 kbp uzunluktadır. Virus, maymun hücrelerinde CPE meydana getirerek litik bir üreme tarzına sahip olmasına karşın, rodent hücrelerinde transformasyonlar yapabilir. Viral genom başlıca 3 bölgeden oluşmaktadır (replikasyon orijini, erken protein gen bölgesi ve geç protein gen bölgesi). Erken gen bölgesi, virusun replikasyonunda ve transformasyon oluşturmasında rolü olan büyük (T) ve küçük (t) ile ifade edilen proteinlerin ve geç gen bölgesi de kapsid proteinlerini sentezlemede ve paketlemede fonksiyona sahiptirler.
SV40’dan bir çok tarzda rekombinant viral vektör hazırlanmış ve çeşitli genlerin transferinde de kullanılmıştır. Hazırlanan başlıca SV40 vektörleri kısaca şöyledir.
a) Virusun erken gen bölgesine yabancı bir gen (Örn., beta globin, Influenza virusu HA geni, vs.) birleştirilirse, bu bölge inaktive olur ve SV40 defektif hale gelir. Böyle vektör, hücrelere transfekte edildiğinde, hücre içinde T-proteini (antijeni) sentezlenemediğinden, virus replike olamaz ve transformasyonlar da yapamaz. Bu vektörün replikasyonu için, hücre içinde sağlam (veya erken gen bölgesi sağlam) SV40 virusu (yardımcı virus, helper virus) bulunan COS hücrelerine (maymun hücresi) aktarılması gereklidir. Defektif viral vektör bu hücrelerde, kendisinin replikasyonu için lüzumlu olan T-antijenini bularak replike olmaya başlar ve yabancı gen ile birlikte kapsid içine paketlenerek olgun viruslar meydana gelir.
b) Eğer, yabancı gen, SV4O’ın geç gen bölgesine aktarılırsa bu bölge inaktive olur. Virus, normal hücrelere transfekte edildiğinde, kapsid sentezi ve paketleme olamayacağından olgun virus teşekkül edemez. Bu olumsuzluğu gidermek için, defektif virus, COS hücrelerine aktarılır. Bu hücrelerin içinde bulunan normal SV40 virusu tarafından sentezlenen, fakat kendisinin kodlayamadığı kapsid içine girer ve olgun virus meydana gelir. Böylece yabancı gen olgun viruslara ve bunlar aracılığı ile de, hücrelere aktarılarak ekspresyonları sağlanmış olur.
SV40 virusu ve gen transferi yandaki şekilde gösterilmektedir. | 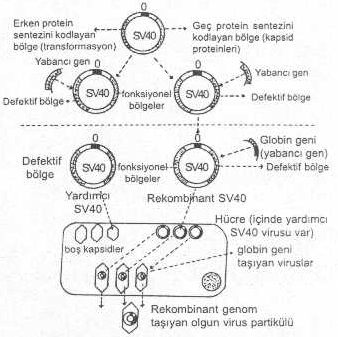
|
c) Eğer, SV40’ın erken gen bölgesi yabancı bir genle inaktive edildikten sonra, içinde, her iki gen bölgesi aktif ve fakat replikasyon orijin bölgesi çıkarılmış mutant bir SV40’la infekte edilmiş hücrelere aktarılırsa (bu mutant virus hücrelerde replike olamamasına karşın, hücrelerin kromozomu ile birleşebilir ve T-antijeninin sentezini uyarır) defektif virus hücrede T-proteinlerini bulacağından replike olur ve kapsid içine paketlenebilir.
d) SV40’dan plasmid tabanlı vektörler de hazırlanmış ve bir çok genin ekspresyonunda da kullanılmıştır. Yabancı gen, önce bir plasmide (pBR322, vs.) aktarılır (plasmidin tetr geni içine integre edilerek bu gen inaktive edilir). Plasmid böylece replikasyon orijin bölgesi ampr geni ve yabancı gen sekanslarından oluşmuş olur. Eğer istenirse, bu aşamada, plasmid bir bakteriye (E. coli ’ye) aktarılarak genin fazla üretimi sağlanabilir. Bakteri lize edildikten sonra, hem plasmid ve hem de plasmiddeki yabancı gen çıkartılır ve viral vektöre bağlıdır. Bu plasmide, SV40’ın promotoru ile regulatör gen sekansları da eklenir. Ancak, yabancı gen, SV4O’a ait sekanslar arasında bulunmalıdır. Böylece plasmid tabanlı rekombinant viral vektör (Örn. pSV2) elde edilmiş olur. Bu vektör, içinde SV40 virusu bulunan yardımcı hücrelere transfekte edildiğinde, plasmidde bulunan SV40 ait sekanslarla, hücre içindeki virusa ait özdeş sekanslar arasında homolog rekombinasyonla birleşme meydana gelir ve yabancı gen, hücredeki viral genoma aktarılır. Bununla birlikte kapsid içine paketlenir .
SV 40’la hazırlanan plazmid tabanlı vektör yandaki şekilde gösterilmektedir. | 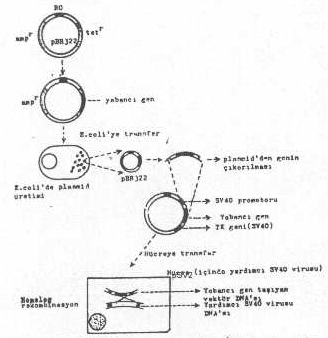
|
e) Yabancı gen (beta globin, vs.) içeren plasmide (pBR322), SV4O’ın sadece replikasyon orijin bölgesi sekanslarına bağlandıktan sonra içinde T-antijeni sentezlenen COS hücrelerine aktarılırsa, vektörün çok sayıda kopyası ile birlikte globin mRNA’sı da meydana gelir ve gen eksprese olur.
f) Yabancı gen (HSV-Tk geni) taşıyan pBR322’ye SV40’ın sadece replikasyon orijin bölgesi de bağlandıktan sonra elde edilen bu rekombinant vektör, Tk- fare hücrelerine aktarılırsa, bu hücreleri Tk+ hale getirirler. Vektör bu hücrelerin kromozomları ile birleşir ancak replike olamaz. Eğer, böyle hücreler, içinde T-antijeni sentezlenen hücrelerle füsyona tabi tutulursa, oluşan hibrid hücrelerde üretilen T-antijenleri Tk+ hücrelerin çekirdeklerine girer, kromozomla birleşir ve vektörün bidireksiyonal replikasyonuna yol açar. Böylece, hibrid hücre içinde çok sayıda Tk+ geni taşıyan sirküler vektör elde edilir (SV40 orijini+HSV-Tk+amp+).
Bu plasmid tabanlı vektör E. coli ’de de replike olabilir ve HSV-Tk geni pozitif koloniler elde edilebilir (mekik vektör).
Yabancı gen, prokaryotiklere ait bir gen (guanosin fosforibosil transferase enzimi, gpt) ise ve bu gen, içinde SV40’ın sadece replikasyon orijin bölgesi promotoru bulunan bir plasmide (pBR322) aktarılırsa oluşan vektör (pSV2gpt) mekik vektör olarak kullanılabilir.
SV40’dan hazırlanan çeşitli vektörlerle, beta-globin, HBsAg, Adv EI A proteini, İnfluenzae virusu-Ha ve NS genleri, vs. hücrelere aktarılarak ekspresyonları sağlanmıştır.
2) Vaccinia virus vektörleri: Vektör olarak hazırlanan ve fazlaca kullanılan viruslar arasında Vacciniada bulunmaktadır. Virus DNA karakterinde bir genoma sahip olup çift iplikcikli, lineer ve yaklaşık 187 kbp uzunluğundadır. Viral genom 150-200 gen ürünü kodlayabilecek bir kapasitededir. Geniş bir hayvan türünde infeksiyon oluşturabilir ve çeşitli hücrelerde de üreyebilir. Hücre içinde, infeksiyondan 12-15 saat sonra 15000 den fazla virus kopyasına rastlanabilir. Vaccinia virusuna eklenecek sekansların üst sınırı henüz saptanamamış olmasına karşın 25 kbp uzunlukta yabancı bir geni kabul edebilmektedir. Vaccinia virusunda bir çok, markerler (Tk, çeşitli temperatür sensitivite ve konakcı genişliğini belirleyen sekanslar, vs.) bulunmaktadır.
Vaccinia virusundan değişik şekilde vektörler hazırlanmış ve kullanılmıştır.
a) Vaccinia virus genomunda bulunan Timidin kinase (Tk) geni, virusun hücre içi replikasyonu için pek gerekli değildir ve infektiviteye de bir katkısı bulunmamaktadır. Dolayısıyla, bu gen içine yabancı DNA sekansları integre edilebilir ve vektör olarak kullanılabilir. Ancak, aktarılan genin ekspresyonu zayıf olur.
b) Vaccinia virusundan, aynen SV40 da olduğu gibi, plasmid tabanlı rekombinant vektörler hazırlanabilir. Burada da yardımcı virus ve hücrelerden yararlanılır.
Vaccinia virusu ve gen transferi yandaki şekilde gösterilmektedir. | 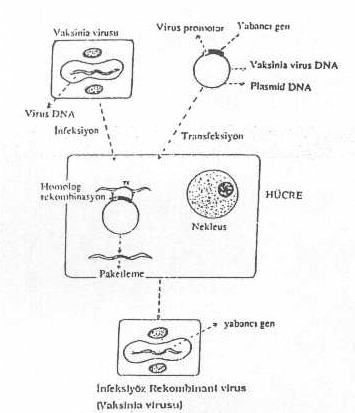
|
Vaccinia virusu vektörleri bir çok genin (HSV-Tk, Influenza virus-HA, Hepatitis B yüzey antijeni, HSV-glikoprotein D, kuduz virusu glikoproteini, VS-G proteini, IB-spike geni, vs.) aktarılmasında vektör olarak kullanılmıştır.
Rekombinant vaccinia virus vektörleri, gen transferleri yanı sıra, biyolojik aktif moleküller üretmek, rekombinant aşılar hazırlamak, yabancı genleri hücre içinde izlemek, gen replasman sağaltım çalışmaları yapmak, vs. amaçlar için de kullanılmaktadır.
Vaccinia genomuna birden fazla yabancı gen integre edilebilmekte ve bunların hücre içi ekspresyonları sonunda, gen ürünleri sentezlenebilmektedir. Örn., vaccinia virusuna, IB+Marek+NDV’larının kapsidlerini kodlayan genler promotorlar ile birlikte integre edilebilir ve böylece polivalan aşılar hazırlanabilir.
3) Polyoma virus vektörleri: Papova grubunda bulunan polyoma virusu, 5.2 kbp uzunlukta, çift iplikcikli, süpersarmal bir DNA’ya sahiptir. Fare hücrelerinde litik üreme tarzı gösterir. Bu özelliği ile SV4O’a benzerliği vardır. Bundan da, infeksiyöz olan ve olmayan vektörler hazırlanmış ve polyoma yardımcı hücrelerinde (MOP) üretilmişlerdir. Polyoma virus vektörlerine, fare İmmunglobulin A zinciri geni aktarılarak elde edilen vektör fare myeloma hücrelerine transfekte edilmiş ve infeksiyondan 48-72 saat sonra, hücrelerde IgA sentezlenebildiği saptanmıştır.
4) Adenovirus vektörleri: İnsanlara ait Adv-2 ve -5 virusları bazı genlerin transferlerinde kullanılmıştır. Viruslarda DNA karakterindeki genomlar lineer olup yaklaşık 35 kb uzunluktadır. Viral DNA’nın iki ucunda tersine tekrarlanan terminal sekanslar bulunmaktadır. Adenoviruslardan, yardımcı viruslara bağımlı olmayan (helper indipendent) vektörler gen aktarımlarında ve genlerin ekspresyonlarında denenmişlerdir. Adenoviruslarında, non-essential coding olan E3 bölgesi içine yabancı gen katılarak E3 promotorunun kontrolü altında, veya E4 bölgesi ile tersine tekrarlanan terminal sekansların (kodlama yapmayan bölge) arasında Adv-major promotorunun kontrolü altında, yabancı genler integre edilerek hazırlanan vektörler, hücrelere aktarılarak ekspresyonları sağlanmıştır.
Adv-5’e, HSV-l glikoprotein B geni bağlanarak hücrelerde ekspresyonu yapılmış ve fareleri virusun letal dozuna karşı koruduğu belirlenmiştir. Adv-2 ve -5’ten infeksiyöz olan ve olmayan vektörler hazırlanmış ve HBsAg, vs. genlerin klonlanmasında ve ekspresyonunda kullanılmıştır.
5) Herpes virus vektörleri: Bu grupla bulunan virusların DNA karakterindeki genomu lineer olup yaklaşık 150 kbp uzunluğundadır. Konakcı hücrelerinde, genellikle, latent infeksiyonlar oluşturur. İnsanlara ait olan HSV1 ve Varicella zoster viruslar (VZV) nöronlarda latent infeksiyon ve Epstein-Barr virusu (EBV) da hem beta -ve hem de tükürük bezi hücrelerinde benzer tarzda üreme karakteri gösterirler. Kanatlıların Marek Hastalığı virusu (MDV) ve hindilerin herpes virusları (THV) da bu grupta olup kanatlıları aşılamalarda aşı virusu olarak denenmişlerdir.
HSV-1 virusundan plasmid tabanlı ve HBsAg yabancı geni taşıyan rekombinant vektör hazırlanarak, içinde yardımcı HSV üreyen hücrelere transfekte edilmiş ve homolog rekombinasyonla yabancı gen, hücre içindeki virus genomuna aktarılarak genin ekspresyonu sağlanabilmiştir. Viruslarda Timidin kinaz (Tk) geni, virusun hücre içinde replikasyonunda gerekli olmadığı (nonessential) için yabancı genler, genellikle, Tk geni içine integre edilmektedir. Bunun dışında, HSV’lerin % 20 kadar bölgesinin nonessential olduğu ve virusun infektivitesinde rolünün olmadığı da belirtilmektedir. Herpes viruslarının çift iplikcikli büyük bir genoma sahip olmaları, bunların, Vaccinia virusları gibi birden fazla geni taşıyabilecekleri (farklı serotiplere ait veya farklı cinslere ait etkenlerin genleri) açıklanmaktadır. Herpes virusları yaklaşık 10 kb uzunluğunda yabancı geni tolere edebilmektedir.
Kanatlı aşılarının elde edilmesinde de, gerek MDV ve özellikle, THV vektör olarak kullanılabileceği belirtilmiştir.
6) Bovine papilloma virusu tip-1 (BPV-1) vektörleri: Bu virus, in vivo olarak keratinositlerde ve in vitro olarak ta konakçı hücrelerinde üreyebilmektedir. BPV-1, DNA karakterinde çift iplikcikli sirküler bir genoma sahip olup yaklaşık 8 kbp uzunluktadır. Dar bir hücre spektrumu göstermesine karşın, bazı fare cell-line’lerinde malignansi oluşturan virus, bir çok genin (insan interferon B1, HBsAg, SV40‘ın erken gen bölgesi, rat insülin geni. vs.) fare hücrelerine (C127, 1 D 13, rat FR 3T3, vs.) aktarılmasında etkin bir vektör olarak kullanılmıştır.
BPV-1 den değişik uzunlukta (3.10, 3.7, 4.4 kb. vs.) plasmid tabanlı rekombinant vektörler (pBPV-1 8-2, pBPV-1 9-1, pBPV-1 69T vs.) hazırlanarak, içinde helper virus bulunan hücrelere transfekte edilmiş ve ekspresyonu sağlanmıştır.
Plasmid tabanlı BPV-1’den mekik vektörler de hazırlanmıştır.
7) Epstein Barr virus vektörleri: Bu virus da klonlamada vektör olarak denenmiştir.
04.02. RNA Virus Vektörleri
RNA karakterinde genoma sahip viruslardan bazıları, özellikle, retroviruslar rekombinant vektör olarak fazlaca kullanılmaktadırlar. Hücrelerin kromozomları ile birleşme özelliği taşıyan retrovirusların replikasyon tarzları, diğer viruslarınkinden farklıdır. Lineer tek iplikcikli (iki moleküllü) 5-7 kb uzunlukla bir genetik materyale sahip olan virus, hücre içine girince sitoplasmada, revers transkriptase enzimi yardımı ile, genomik RNA’ya tek iplikçik cDNA (copy DNA, komplementer DNA) sentezlenir. Sonra, bu cDNA’ya da polimeraz 1 enzimi ile ikinci bir cDNA sentezlenerek çift iplikcikli DNA’ya dönüştürülür (pro virus). Bu pro virus, hücre çekirdeği içine girerek kromozomla birleşir ve transkripte olur. Oluşan viral RNA sitoplasmaya geçer ve burada sentezlenen kapsid içine paketlenir. Olgun virus hücrelerden tomurcuklanarak dışarı çıkarlar.
Vektör olarak yararlanılan çift iplikcikli sirküler pro virusun her iki ucunda long terminal repeat (LTR) olarak tanımlanan ve yüzlerce baz çiftinden oluşmuş tekrarlanan uzun sekanslar bulunmaktadır. Bu uçlar transkripsiyonu başlatmada ve bitirmede etkin bir regulatör sinyal mekanizması görevine sahiptirler (promotor). Provirus, LTR’lerden sonra, zarf proteini (env), RNA’ya bağımlı DNA polimeraz (reverse transcryptase, pol) ve grup antijenleri (gag) gibi bölgelere de sahiptir. Yabancı gen, provirusun gag, env, vs, gen bölgelerinden biri içine integre edilir ve hücrelere (NIH 3T3, vs.) transfekte edilir (helper virus içeren yardımcı hücreler). Vektörün hücre içinde replikasyonu ve genin ekspresyonu sağlanır.
Retrovirus genomu ve vektörü yandaki şekilde gösterilmektedir. | 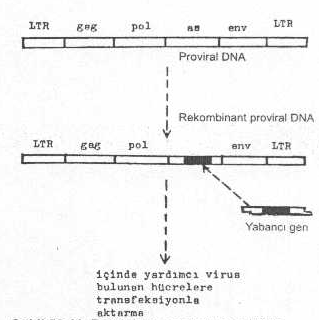
|
Retroviruslarından, ayrıca, plasmid tabanlı vektörler hazırlanmış ve gen aktarımında kullanılmıştır. Retroviruslardan olan kanatlıların avian leukosis virusu (ALV) ve avian reticuloendotheliosis virus (REV) viruslarından özellikle ikincisi, onkojenik olmaması nedeniyle vektör hazırlanmasında tercih edilmektedir.
Baculovirus Vektörleri
İnsektlere ait olan Baculoviruslar da bazı genlerin (kanatlıların Newcastle Hastalığı virusuna ait bazı genler) aktarılmasında vektör olarak denenmişlerdir
05. Bakteriyel Vektörler
Yabancı genleri klonlamada, her ne kadar virus ve plasmidler kadar yoğun bir kullanım alanı bulamamasına karşın, bakteriyel vektörler son yıllarda bazı genleri bakteri genomlarına klonlamada başarı ile denenmiştir. Bunlar arasında başlıca,
S. typhimurium ’un okzotrofik bir mutantı olan AroA SL 3261 suşu (üremesi için p-aminobenzoata gereksinimi vardır), K88 fimbria, termolabil-B toksini ve 65 kD’luk mikobakteri (M. tuberculosis) antijenik determinantlarını klonlamada kullanılmıştır. Bu tarzda hazırlanan rekombinant bakteriyel vektörler, farelere verildiğinde hem S. typhimurium ’a ve hem de taşıdığı gen ürünlerine karşı humoral ve sellüler bir yanıtın meydana geldiği açıklanmıştır.
S. typhi ’nin genetik modifiye mutantı olan S. typhi Ty2la suşu (galactose epimerase negatif), S. sonnei antijenik determinantlarının klonlanmasında kullanılmıştır.
E. coli K12 suşu da V. cholerae antijeni için aynı amaçlarla denenmiştir.
Bacille Calmette Guerin (BCG) suşuna da yabancı genler aktarılarak bunların ekspresyonunun sağlanması olanakları araştırılmakta ve ayrıca BCG’nin adjuvant etkisinden de yararlar sağlanmaktadır.
[1] Kaynak : Temel Mikrobiyoloji