Yer Değiştirebilen Genetik Elementler
Prof. Dr. Mustafa Arda
Ankara Üniversitesi Veteriner Fakültesi
Temel Mikrobiyoloji 1
01. Giriş
02. İnsersiyon Sekansları (İS-Elementleri)
03. Transposonlar (Tn)
03.01. Replikatif Transpozisyon
03.02. Nonreplikatif (Direkt) Transpozisyon
03.03. Konservatif Transpozisyon
04. Mu Fajı (Mutator Faj)
05. İntegronlar
06. Önemli Notlar
01. Giriş
Yer değiştirebilen genetik elementlere (Transposable genetik elementler) prokaryotik ve ökaryotik organizmalarda fazlaca rastlanmaktadır. Bakteri, mantar, maya, mısır, drosofila, vs. bunlardan sadece birkaçıdır. Bu elementler, bir kromozomdan diğerine, aynı kromozom üzerinde bir bölgeden başka bir yere, kromozom ile plasmid veya iki plasmid arasında yer değiştirebilen, kendinin transferini sağlayabilen küçük, lineer ve çift iplikçikli DNA sekanslarıdır. Plasmidlerde rastlanan transposable elementler, aynen kromozomda olduğu gibi, bir plasmid üzerinde pozisyonunu değiştirebilir veya iki plasmidi birbirine birleştirebilir. Transposon ile hedef bölge sekansları arasında (girdiği yeni bölge) homolog yanların bulunmasına gereksinim yoktur.
Transposable elementler (mobil elementler) ilk defa, 1940 yılında Barbara McClintock tarafından mısır bitkisi genetiği üzerinde yaptığı çalışmalar sırasında saptanmıştır. Sonradan, bakteri, maya, diğer bazı ökaryotiklerde de varlığı bildirilmiştir.
Bakterilerin yeni karakterler kazanmasında ve yeni mutasyonların oluşmasında etkinlikleri fazla olan transposable elementler başlıca 3 grup altında toplanmaktadırlar.
1) İnsersiyon sekansları (İS-elementleri)
2) Transposonlar (Tn)
3) Mu fajı (mutator faj, mutasyon oluşturan faj)
02. İnsersiyon Sekansları (İS-Elementleri)
Transposonlardan daha basit bir yapı karakteri gösteren İnsersiyon sekansları (İS-elementleri) yaklaşık 750-1500 bp uzunluktadırlar. Elementlerin türlerine göre değişmek üzere, iki ucunda 10-40 bp kadar olan ve tersine tekrarlanan sekanslar (inverted repeat,IR) bulunmaktadır. Bunları dış taraftan çevreleyen, yeni girdiği alıcıya ait hedef bölgede ve aynı oriyentasyonda, 5-12 bp uzunlukta direkt tekrarlanan (direct repeat, DR) nukleotid sekansları yer almaktadır. İS-elementlerinin iki ucunda lokalize olan tersine tekrarların arasında, bu elementlerin DNA üzerinde yer değiştirmesinde rolü olan ve elementlerin sınırlarını iyi belirleyen transpozisyon enzimini kodlayan gen (transposase) bulunur.
Yandaki şekillerde bir İS-elementinin , bir transposonun ve bir kompozit transposonun yapısı gösterilmektedir. | 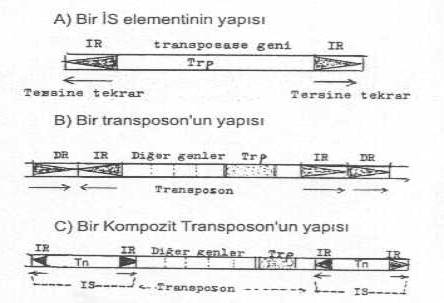
|
Aşağıdaki tabloda E. coli 'ye ait bazı İS-elementleri ve özellikleri gösterilmektedir.
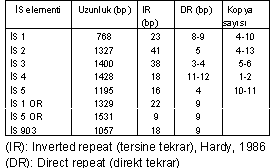
İS-elementlerinin lokasyonlarını, kromozom veya plasmid üzerinde belirlemek oldukça zordur. Ancak, genin içine girdiğinde geni inaktive edebilir, fonksiyonunu bozabilir ve transkripsiyon terminasyon sinyali taşıdığında, genlerin transkripsiyonuna mani olabilirler.
E. coli kromozomunda ve plasmidlerinde değişik yerlere lokalize olmuş birçok İS-elementlerine rastlanmıştır. Bunlar, bakteride, olumlu veya olumsuz sonuçlar oluşturan değişikliklere yol açmaktadırlar. Özellikle, İS-elementleri arasında rekombinasyonların meydana gelmesi ve bir plasmiddeki İS-elementinin kromozom veya diğer bir plasmide transfer olması böyle değişiklikleri daha da arttırmaktadır.
İS-elementi, DNA üzerinde eski yerinden çıkarak yeni bir hedef bölgeye girdiğinde, alıcının bu bölgesinde bulunan kısa sekanslar (hedef sekanslar) dublike olurlar. Bu bazlar direkt tekrar (DR) karakteri taşırlar. Çünkü, aynı oriyentasyona sahiptirler. Başlangıçta transpozisyon olmadan önce, hedef bölgede sadece tek bir çift iplikçikli sekans bulunurken, insersiyon olduktan sonra, bu sekanslarda dublikasyon meydana gelerek, İS-elementinin iki ucunda, IR sekanslarına dıştan birleşmiş olarak, bir lokalizasyon gösterirler.
Bir İS-elementinin hedef DNA’da lokalizayonu yandaki şekilde gösterilmektedir. | 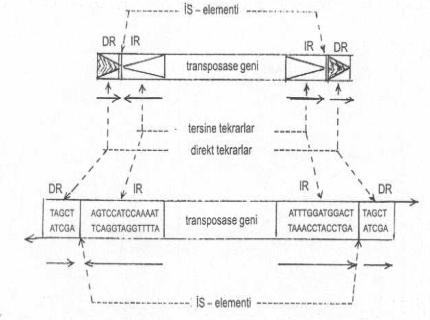
|
Konakçıda hedef bölgeye ait baz sıralarının sayısı ve nukleotidlerin türü, İS-elementinin (veya transposonun) karakteri ve türü ile ilişkilidir.
Alttaki şekilde, konakçı DNA'sında bulunan hedef bölgeye ait İS-elementinin (veya transposonun) girmesi ve hedef bölge sekanslarının dublike olması ve elementin iki ucunda direkt tekrar olarak yer alması gösterilmektedir.
Bir İS-elementinin hedef DNA bölgesine insersiyonu yandaki şekilde gösterilmektedir. | 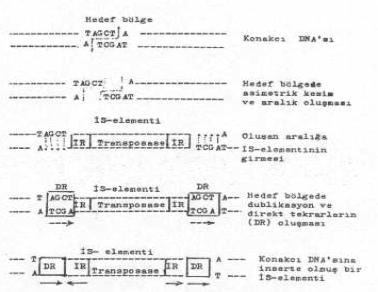
|
İS-elementleri, transposonların iki ucunda da bulunabilirler. İS-elementlerine plasmidlerde de fazlaca rastlanılmaktadır.
Yandaki şekilde bir R plasmidinde bulunan transposon ve İS-elementleri gösterilmektedir. | 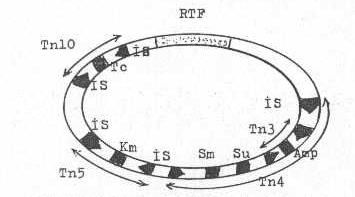
|
İS-elementlerinin, transposonlar için, DNA üzerinde hareket edebilen hedef sekanslar oluşturmada da etkinlikleri vardır (mobil hedef bölgeler). Ayrıca, İS-elementleri, kompozit transposonlarının sonlarını da belirlerler.
03. Transposonlar (Tn)
Transposonlar, İS-elementlerine oranla, daha büyük ve komplike bir özellik gösterirler. Yapılarında, transposase geninden ayrı olarak özel marker genleri (antibiyotiklere, kemoterapötiklere, metallere dirençlilik, vs) taşırlar. Transposonların yapısı, temel karakterleri bakımından, İS-elementlerine benzerlik gösterir. Transposonlar iki ucunda tersine tekrar (IR), bunları dıştan çevreleyen direkt tekrar (DR) ve ortada da transposase geni ayrıca başka diğer özel marker genleri de bulunmaktadır.
Eğer transposonların iki ucunda İS-elementi (İS modülü) bulunuyorsa bunlara genel olarak kompozit transposonlar adı verilir. Eğer, iki uçta, IR ve DR sekansları varsa bunlara basit transposonlar (Tn3 benzeri transposonlar) denilmektedir.
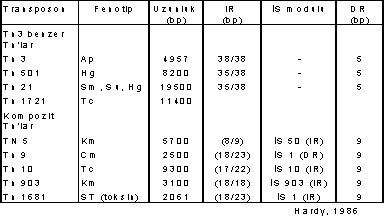
Aşağıdaki çizelgede E. coli 'ye ait transposonlar ile bunların bazı özellikleri gösterilmektedir.
İS-elementleri ve transposonlar birer replikon değildirler. Yani, bakteri içinde bağımsız olarak replike olamazlar. Çünkü, replikasyon orijini sekanslarına sahip değildirler. Bakteri ve plasmid DNA'sı dışında da bir etkinlikleri yoktur. Aynı durum ökaryotiklerdekiler için de geçerlidir.
Transposonlar ve İS-elementleri genom üzerinde veya plasmidlerde devamlı yer değiştirdiklerinden mutasyonlara da neden olmaktadırlar. Genlerin içine veya aralarına, bu elementlere ait kısa veya uzun sekansların (1000-2000 bp) girmesi transkripsiyon ve translasyonun bozulmasına yol açtığı gibi, baz sıralarının değişmesine ve buna bağlı olarak ta oluşan yeni tripletler (kodonlar) mutasyonların meydana gelmesine sebep olurlar. Bu yeni kodonlar, mRNA'nın da değişmesine ve yeni karakterde tripletlerden oluşmuş mRNA'ya ve değişik proteinlerin oluşmasına yol açarlar. Bu proteinler etkisiz olabileceği gibi farklı yapıda da olabilirler. Bazı durumlarda da, yeniden stop veya terminatör sekanslar mRNA'ya girerek tam olmayan peptidlerin meydana gelmesine neden olurlar. Transkripsiyonun durmasına yol açarlar.
Bazen de, transposable elementler, konakçı genomuna yeni promotorlar katarak genlerin aktivasyonuna ve bazı transposonlar da, genlerin içine girdiklerinde genlerin bütünlüğünü bozarak genin inaktivasyona neden olurlar. Böyle durumlar, eğer bakteri için çok önemli olan ürünün veya proteinin sentezini inhibe etmişse, ölümlerde meydana gelebilir.
Transposonlar, üzerinde bulunduğu genomda yeni rekombinasyonlar için hareketli hedef bölgeler oluştururlar. Böylece, aynı veya ayrı yerlerde, biri bakteride ve diğeri de plasmidde, veya iki ayrı uygun hedef bölgeler arasında, karşılıklı rekombinasyonlar için uygun bölgeler sağlandığı gibi yer değiştirme olanağı da meydana getirilir. Plasmidlerde bulunan transposonlar, plasmidin kromozomla da birleşmesine neden olabilmektedirler.
Transposonlar, aynı zamanda plasmidle konakçı kromozomu arasında da hareket ettiklerinden, tek veya çoğul rezistenslik genlerini de birbirlerine aktarabilirler. Böylece, rezistenslik genlerinin yayılmasına neden olurlar.
Transposonlar, çoğu antibiyotiklere ve kimyasal maddelere karşı dirençlilik genleri taşıdıklarından, bir plasmid veya kromozomda bulunan Transposonlar mikroorganizmalara tek veya multiple dirençlilik kazandırırlar.
Transposonların yer değiştirmesi sırasında kopyalarının meydana gelmesi ve bunların kromozom üzerinde değişik bölgelere girmesi sonu oluşan mutasyonların yarattığı olumsuzluk kadar, transposonların DNA üzerinden ayrılması da aynı tarzda benzer sonuçlar meydana getirir.
Transposonların genom üzerinde hareketleri nedeniyle, bazı genler inaktive olurken (kapatılırken), kapalı olanlar da açılabilirler (aktive olabilirler).
Bakteriyel transposonlar, genom üzerinde veya genom ile plasmid arasında kendi transpozisyonunu sağlayabilecek gerekli bilgilere sahipse de, yine de üzerinde bulunduğu konakçının yardımcı faktörlerine, aktivitesine ve enzimlerine (DNA pol, DNA gyrase, vs) ihtiyaç gösterirler.
Kompozit Transposonların sonlarında bulunan İS-elementleri (İS modülü), bazılarında direkt tekrar (DR) tarzındadır (Tn 9 da olduğu gibi). Bir kısmında da tersine tekrar (IR) halindedir (Tn 5, Tn 10, Tn 903, Tn 1681).
İS-elementlerinde ve transposonlarda bulunan transposase enzimi hem hedef bölgeyi tanımada, elementlerin sonlarını belirlemede ve hem de yer değiştirmede etkin role sahiptir.
Transpozisyon başlıca 3 mekanizma ile gerçekleşmektedir. Bunlar da,
03.01. Replikatif Transpozisyon
Replikatif transpozisyon, DNA üzerinde bulunan transposonda bir çoğalmanın (dublikasyon) meydana gelmesi ve orijinal kopyanın parental DNA'da kalarak, yeni sentezlenen kopyanın başka bir hedef bölgeye transfer edilmesi tarzında gerçekleşen bir yer değiştirme olgusudur. Bu tür transpozisyonda transposase enzimi yanı sıra resolvase enzimi de yardımcı olur. Bu son enzim, yeni kopyanın hedef bölgeye girmesinde (rekombinasyonda) etkinlik gösterir.
Replikatif transpozisyonun aşamaları yandaki şekilde gösterilmektedir. | 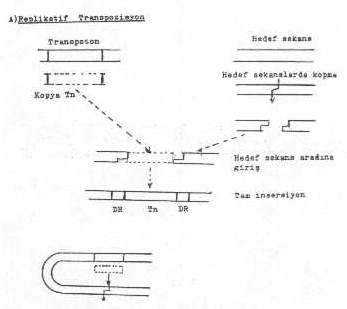
|
Transpozisyon için, konak hücre kromozomu ile hedef bölge arasında homolog sekanslara gerek yoktur.
03.02. Nonreplikatif (Direkt) Transpozisyon
Transposon, bulunduğu yerden çıkarak,yeni hedef bölgeye direkt olarak geçebilir ve bu bölge içine girebilir. Bunun için, önceki bulunduğu DNA'da transposonun iki ucunda kesimler meydana gelerek, Tn buradan ayrılır, bunun karşısına gelen yeni hedef bölge sekansları arasında benzer tarzda oluşan kesimler sonu aralanan DNA içine girer. Bu tür transpozisyona Tn 5 ve Tn 10 da rastlanmaktadır. Direkt transpozisyonda transposase enzimi etkilidir. Verici DNA'dan çıkan Transposonun geride kalan açıklığı hemen kapatılır. Eğer hücre bunu kapatmazsa, ölümler meydana gelebilir.
Nonreplikatif transpozisyonun aşamaları yandaki şekilde gösterilmektedir. | 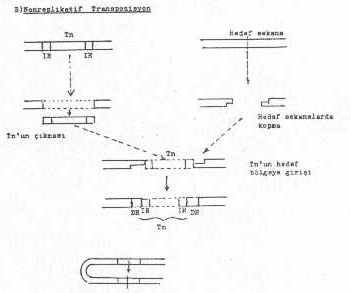
|
03.03. Konservatif Transpozisyon
Bu tür transpozisyon lambda fajında olduğu gibi gerçekleştirilir. Karşılıklı gelen sekanslar arasında çapraz rekombinasyonla integrasyon meydana gelir.
Bazı transposonlar sadece bir tür ve bir kısmı da birden fazla mekanizmayı kullandıkları saptanmıştır. Örn, İS1 ve İS 903 hem replikatif ve hem de nonreplikatif mekanizmayı kullanabilmektedir.
Transposonların ve İS-elementlerinin yer değiştirme mekanizmaları oldukça komplike bir olgudur. Yeni bölgeye girmede meydana gelen rekombinasyon hem direkt ve hem de tersine tekrarlar arasında oluşabilir.
Bazı durumlarda da kromozom veya plasmidler üzerinde bulunan direkt tekrarlar veya tersine tekrarlar arasında karşılıklı rekombinasyonlar da meydana gelebilmektedir. Böylece karşılıklı genetik materyaller de transfer edilmiş olur.
Yandaki şekillerde tekrarlar arasındaki rekombinasyonlar gösterilmektedir. | 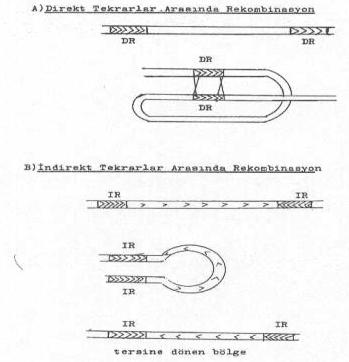
|
Transposon taşıyan bir replikon (Tn+) ile taşımayan (Tn-) diğer bir replikon yan yana geldiklerinde ve birleşme olduktan sonra iki tane transposon taşıyan kointegre büyük bir molekül ve bundan da birer kopya Tn taşıyan iki replikon meydana gelir.
Transposon birleşmeleri ve kointegrasyon oluşumu yandaki şekilde gösterilmektedir. | 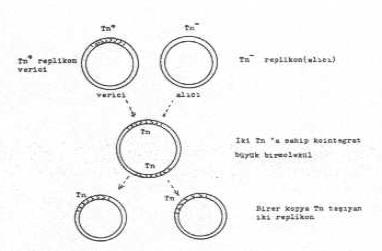
|
4. Mu Fajı (Mutator Faj)
E. coli 'ye ait olan bu faj, aynı bir transposon gibi bakterinin kromozomu üzerinde yer değiştirebilir ve çeşitli yerlere girebilir. Böyle durumlar mutasyonlara yol açmaktadır. Bu faja aynı zamanda "dev transposon" adı da verilmektedir. Mu fajı, E. coli 'de mutasyon meydana getirmek için kullanılmaktadır. Mu fajı, konakçı bakteriyi infekte ettikten sonra, genoma nonreplikatif tarzda integre olduğu açıklanmaktadır. Litik dönemde fajın, replikatif transpozisyonla da bir çok kopyası çıkmaktadır.
05. İntegronlar
Son yıllarda, klinik materyallerden izole edilen, Enterobacteriacea familyasına ve pseudomonaslara ait mikroorganizmalarda, bir çok rezistenslik genleri taşıyan ve İntegron olarak tanımlanan yeni hareketli elementlerin (mobile element) varlığı bildirilmiştir. Bunlara, ya bağımsız olarak plasmidlerde veya Transposon (Tn21) ailesinin bir bölümü olarak rastlanılmaktadır. İntegronlarda, temperate fajlarınkine benzeyen integrase geni (int) ve bağlanmada rol oynayan diğer bir genden oluşan site specific recombination sistemi bulunmaktadır. Hemen her integronda, sulfonamid rezistenslik geni (sul I) ve ayrıca bir çoklarında da diğer rezistenslik genlerinin (streptomycin/spectinomycin, aminoglycoside, b -lactamase, trimethoprim, chloramphenicol) varlığı bildirilmiştir.
İntegronların da bakterilere rezistenslik kazandırmada önemli rolleri olduğu belirtilmektedir. Yapılan, PCR ’a dayalı çalışmalarda, integronların DNA haritaları da çıkarılmıştır.
06. Önemli Notlar
1- Bazı yazarlar insersiyon sekanslarını sadece iki uçtaki IR ile sonlandırmakta bazıları da buna her iki tarafında olmak üzere direkt tekrarları (DR) ilave etmektedirler.
2- Bu nedenle ilk şekildeki İS elementinin sonunda DR konmamıştır. Çünkü, DR'ler (direkt tekrarlar) alıcı DNA'sına aittir.
[1] Kaynak: Temel Mikrobiyoloji