Ribosomlar;RibosomalRNA'lar(rRNA) ve Transfer RNA(tRNA)
Prof. Dr. Mustafa Arda
Ankara Üniversitesi Veteriner Fakültesi
Temel Mikrobiyoloji 1
01. Ribosomlar ve Ribosomal RNA'lar (rRNA)
01.01. A-Bölgesi (Akseptör Bölge)
01.02. P-Bölgesi (Peptidil Bölge)
01.03. mRNA Bölgesi
01.04. E (exit) Bölgesi
01.05. Peptidil Transferase Bölgesi
01.06. 16S rRNA Bölgesi
01.07. 23S rRNA Bölgesi
01.08. 5S rRNA Bölgesi
02. Transfer RNA (t RNA)
01. Ribosomlar ve Ribosomal RNA'lar (rRNA)
Ribosomlar, hücre içinde amino asitler arasında peptid bağlarının kurulmasını sağlayan, mRNA'ya bağlandıktan sonra bunun üzerinde 5'®3' yönünde hareket ederek protein sentezini gerçekleştiren kompakt, identik, irregüler bir morfolojik profile sahip olan partiküler strüktürlerdir. Yaklaşık, 15-20 nm büyüklükte olan ribosomlar, aktif üremekte olan E. coli 'ler de sayısı 15000-20000 arasında değişebilir. Hücre içinde serbest olarak bulunabilecekleri gibi, sentez sırasında mRNA üzerinde de tesbih gibi diziler halinde de (poliribosom) bir araya gelebilirler. Prokaryotiklerde ribosom'lar 70S (S: Svedberg ünitesi, sedimentasyon konstantı) karakterindedirler (ökaryotiklerde ise 80S'lik ribosom vardır). Birbirinin benzeri şekil ve yapıda olan ribosomların %60-65'ini ribosomal RNA (rRNA) ve %35-40'ında özel proteinler oluşturur ve MA:2.6x106 daltondur.
Prokaryotiklerde ribosomun yapısı ve komponentleri yandaki şekilde gösterilmektedir. | 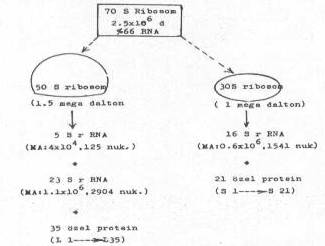
|
Bakterilerin ribosom'ları (70S), biri 30S (MA: 1x106) ve diğeri 50S (MA: 1.6x106) olan iki asimetrik fonksiyonel alt üniteden oluşur: (ökaryotiklerin 80S'lik ribosomu, 40S ve 60S iki alt üniteden meydana gelmiştir ve mitokondriumların ribosomları 50 S 'dir). Bu iki alt ünitenin bağlanmasında divalent katyonlardan olan Mg++ 'nin rolü oldukça fazladır. İki alt ünite birleştiğinde aralarında bir oluk meydana gelir. Ancak, mRNA bu oluğa değil de, 30S'lik alt ünitenin üzerine bağlanır.
E .coli ‘de bir tam ribosomun elektronik mikroskobik görünümü yandaki şekilde gösterilmektedir. | 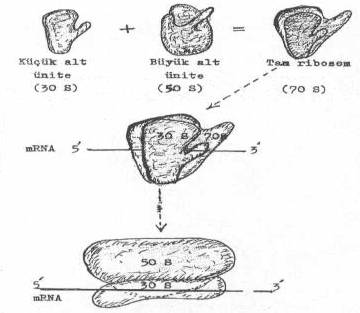
|
Ribosom'ların yapısında 3 tür rRNA bulunmaktadır. Bunlardan 5S rRNA'lar (MA: 4x104, 125 nukleotid) ve 23S rRNA'lar (MA: 1.1x106 2904 nukleotid) 50S'lik büyük alt ünitede, ve 16S rRNA'lar da (MA: 0.6x106, 1542 nukleotid) 30S'lik küçük alt ünitede yer almaktadırlar. Bunlara ilaveten, büyük alt ünitede birbirinden farklı, tek kopya halinde özel 35 protein (L1 ® L35, MA: 10000-20000) ve küçük alt ünitede de 21 tane (S1®S21, MA: 10000-20000) spesifik protein vardır. Bu spesifik ribosomal proteinler elektroforetik olarak, birbirlerinden, molekül ağırlıklarına göre ayrılırlar ve bunların ribosomlar üzerindeki yerleri de oldukça özeldir. Bu proteinlerin bir veya birkaçının çıkarılması ve etkinliğinin olmaması, ribosomların fonksiyonlarına da olumsuz yönde etkiler. Örn., 50S'lik alt üniteden L16'nın çıkarılması peptid formasyonunu bozar. Küçük alt üniteye bağlanan Streptomisin bu ünitenin işlevini inhibe eder. S4 ve S12 proteinlerinde oluşan değişmeler bazı antibiyotiklere karşı duyarlılığı azaltır. Açıklanan nedenler bu proteinlerin, gerek 30S ve gerekse 50S'lik ribosomlarda çok önemli ve özel görevleri olduğunu ortaya koymaktadır. Bu nedenle, proteinlerin ribosomal alt ünitelerindeki yerleri çok spesifiktir.
Ribosomlardan küçük alt ünitenin (30S), mRNA'nın 5' -terminusunda bulunan Shine Dalgarno sekanslarının (SD) ve AUG kodonunun bağlanmasında ve protein sentezinin başlamasında çok önemli etkinliği vardır. Bu alt ünitede bulanan 16SrRNA'nın 3' - terminusundaki baz sıraları aynı zamanda mRNA'nın 5-' ucundaki sekanslara komplementer olması, bu bağlantıyı kuvvetlendirir. Ayrıca, 16SrRNA'da bulunan spesifik bölgeler, direkt olarak, tRNA'nın antikodon bazları ile ve alt ünitelerdeki hem A ve hem de P bölgesindeki nukleotidlerle ilişki kurarlar. Büyük alt ünitede bulunan 23SrRNA ise, peptidli RNA'ların CCA terminusu ile interaksiyonu vardır. Eğer mutasyonlar mRNA ile rRNA arasındaki ilişkiyi veya komplementerliği bozarsa veya zayıflatırsa translasyona da olumsuz yönde etkiler. Colicin E3, 16SrRNA'nın 3' -ucundan 50 'ye yakın nukleotidi çıkarttığından protein sentezinin başlatılması inhibe olur. Ribosomlarda bulunan ve bunların omurgasını oluşturan rRNA'lar ile spesifik proteinler birbirleriyle koordine bir şekilde işbirliği içindedirler. Ribosomların yüzeylerinde ki proteinler de birbirleriyle kimyasal bağlantı kurarlar.
Ribosomlardaki spesifik proteinler, rRNA'lardan daha fazla antijeniktirler. Eğer ribosomlar tümden bir hayvana verilirse kendisine karşı antikor sentezini uyarırlar.
Ribosomlar in vitro koşullarda komponentlerinden ayrılabilirler (59 fraksiyon) ve tekrar birleştirilebilirler. Ancak, bu reasamble işlemi, bir sıra ve düzen içinde gerçekleştirilmesi gereklidir. İki ribosomal alt ünite birbirlerine tek bir proteinle kovalent olarak bağlanmıştır. Ribosomal RNA'lar, tekbir promotordan orijin alarak transkripte edilir ve bundan translasyon gerçekleştirilir.
Büyük ve küçük ribosomal alt ünitelerde birçok aktif yüzeyler bulunur. Bu bölgelerin başlıcaları aşağıda özetle belirtilmiştir.
Bir ribosomun yapısı yandaki şekilde gösterilmektedir. | 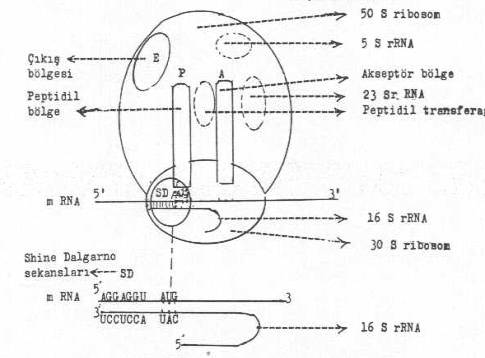
|
01.01. A-Bölgesi (Akseptör Bölge)
Amino asitlerle bağlanmış olan spesifik tRNA'ların antikodonlarının, 30S alt ünitede mRNA üzerinde sırada bulunan bazlarla birleştiği bölgelerdir. Bu bölge hem 50S ve hem de 30S alt ünitede bulunur.
01.02. P-Bölgesi (Peptidil Bölge)
A bölgesinde özel tRNA ile bağlı bulunan aminoasitlerin, bu bölgeyi boşaltmak için aktarıldıkları bölgedir. Bu bölge hem 50S ve hem 30 S'lık alt ünitede bulunur.
01.03. mRNA Bölgesi
Bakterilerde DNA iplikçiklerinin birinden (kalıp ödevi gören ve 3' ¬ 5' yönünde olan) RNA polimerase enziminin katalitik etkisiyle sentezlenen mRNA'nın translasyonu için 30S'lik alt ünitede yerleştiği bölgedir.
01.04. E (exit) Bölgesi
P-bölgesinden ayrılan tRNA'nın 50S'lik alt üniteyi terk etmeden önce, bu alt ünite üzerinde ve geçici olarak bulunduğu bölgedir.
01.05. Peptidil Transferase Bölgesi
A-bölgesi ile P-bölgesi arasında lokalize olan ve peptidil transferase enziminin aktivite gösterdiği ve iki amino asitin birbirine bağlandığı bölgedir.
01.06. 16S rRNA Bölgesi
30S ribosomal küçük alt ünitede bulunan ve 16 S rRNA'nın 3' -terminusundaki sekanslarla mRNA'nın 5' -ucundaki SD sekansları ve AUG kodonları ile bağlanan yerlerdir.
01.07. 23S rRNA Bölgesi
50S ribosomal alt ünitede bulunan A- ve P- bölgelerini kapsayan yerde etkinlik gösteren 23 S rRNA bulunduğu ribosomal alandır.
01.08. 5S rRNA Bölgesi
50 S 'lik ribosom üzerinde bulunur.
02. Transfer RNA (t RNA)
Transfer RNA (t RNA), protein sentezinde çok önemli ve merkezi fonksiyona sahip bir moleküldür. Tek iplikçik ve yaklaşık 75-90 nukleotid'den oluşan t RNA'da, m RNA gibi, RNA polimerase enzimi tarafından DNA'dan sentezlenir. Bütün t RNA'lar (başlatıcı f met t RNA hariç), zincir uzatma faktörü, EF-Tu (elongation factor-Tu) tarafından tanınarak ribosomlara taşınırlar. Buna karşın, f met t RNA ise, başlatma faktörü İF-2 (initiation factor-2) ile birleşerek, ribozomlar üzerindeki AUG kodonuna, kendinde bulunan antikodon bölgesindeki UAC bazılarıyla birleşir.
Her amino asit, birden fazla t RNA ile temsil edilir. Bir amino asiti temsil eden birden fazla t RNA 'ya isoakseptör tRNA adı verilmektedir (Örn, Leucine, 4 amino acyl t RNA: CUU, CUC, CUA ve CUG tarafından temsil edilir). Antikodon civarında meydana gelen değişiklikler, kodon-antikodon birleşmesine de etkilerler.
tRNA üzerinde yapılan çalışmalar, bunun sekonder yapısının 4 kollu, tersiyer yapısının da bir birine dikey L-şeklinde iki heliksten oluştuğunu ortaya koymuştur. Sekonder yapı formunda, tek iplikçikli olan t RNA, kendi üzerine kıvrılarak 4 tane çift iplikçikli kol ve bunların ucunda da tek iplikçik halinde bir ilmek bulunur (akseptör bölge hariç çünkü, bunda ilmek yok). Bir hücrede 50'den fazla farklı t RNA vardır.
Bir tRNA’ nın yapısı yandaki şekilde gösterilmektedir. | 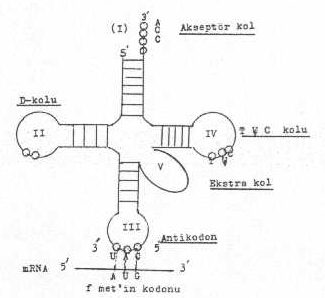
|
1) Akseptör kol (I): Amino asitlerin, kendine ait t RNA'ların bağlandığı serbest uçtur (3' -ucu, CCA). Bu uca, aminoacyl synthetase tarafından tanınarak aktive edilen spesifik amino asitler kovalent olarak bağlanırlar. CCA (adenilat) ucu, bütün t RNA'larda spesifik amino asitlerle ilişki kurabilmektedir (ilmeksiz kol.)
2) D-kolu (II): D-kolu, dihidrouridinden zengindir. Buranın, 7-11 bazlık kısa çift iplikçikten oluşmuş bir bölgesi vardır. Amino acyl synthetase enzimini tanır. Ayrıca, bu bölgenin ucunda tek iplikçikten oluşmuş bir ilmek bölgesi vardır.
3) Antikodon kolu (III): tRNA'da bulunan antikodon bölgesi bazları, mRNA'daki kodon bazları ile anti paraleldir. Bu bölge tek iplikçiktir.
Taşıdıkları amino asitlerin şifrelerine göre antikodondaki baz sıraları değişebilir. Şöyle ki; 30 S ribosomal alt ünite üzerinde, AUG kodonu sırada ise, buna ait metionini buraya taşımak için bu kol met tRNA ile birleşir. Met tRNA'nın da antikodon bölgesinde UAC bazları bulunur. Kodon ve antikodon bazları birbirine anti paraleldir ve kovalent olarak birleşirler.
4) T y C kolu (IV): Bu kısım sıra dışı bazlara sahiptir. Bu kolda pseudouridine (y) ve ribotimidin fazlaca bulunur. Bu kol ribosomları tanır.
5) Ekstra kol: Bu kol, antikodon ile T (y) C arasında yer alır. Bu kol, karakterine göre iki yapısal form gösterir. Biri, küçük ekstra koldur (5-6 bazlık). Yaklaşık bütün t RNA'ların %75'in de vardır. Diğeri ise, büyük ekstra kol olup 13-21 bazdan meydana gelmektedir.
Akseptör kolda 7 baz birleşmesi D-kolunda 3 (bazen 4), antikodonda 5 baz ve Ty C de 5 baz birleşmesi bulunmaktadır. Baz birleşmeleri G-C ve A-U bazları arasındadır. Bazen G-U, G veya A-y da olabilmektedir.
Bu 4 kollu sekonder temel yapı formu, prokaryotik, ökaryotik, mitokodria ve kloroplast tRNA 'larında birbirlerine benzerler.
Transfer RNA, adaptör bir moleküldür. Hücre içinde iki önemli fonksiyonu vardır. Bunlardan biri akseptör kolun 3' -CCA ucuna amino asitlerin kovalent bağlanması ve diğeri de kendinde bulunan antikodon bölgesiyle m RNA üzerindeki kodonla birleşmesidir. Transfer RNA'nın antikodon bölgesinde 3 bazdan oluşan triplet vardır ve bu bazlar kodon bazlarına anti paraleldir. Diğer bir deyimle, kodondaki 1. baz ile antikodondaki 3. baz karşılıklı kovalent bağlarla birleşirler.
Protein sentezinin başlangıcında 70 S 'lik ribosomun 30 S alt ünitesi üzerindeki P-bölgesinde m RNA'nın AUG başlatma kodonu bulunduğunda, hücre içinde ki özel amino acyl synthetase enzimi, bu kodona ait olan formilize olmuş metionini (f met) bularak aktive eder ve kendine ait t RNA'nın 3' -CCA ucuna kovalent olarak bağlanır. Bu, f met t RNA'nın antikodon bölgesinde ise UAC tripleti vardır. f met tRNA, IF-2 ile birleşerek kodona taşınır ve onunla birleştirilir. Bunun dışındaki diğer tRNA 'lar ise EF-Tu ile birleşerek ribosom üzerine gelirler.
Hücre içinde her aminoasit için bir ve birden fazla spesifik tRNA bulunur. Amino asitlerin kendine özgü t RNA'ları ile birleşmesini amino acyl synthetase enzimi katalize eder. Hücre içinde yaklaşık 3000'den fazla her tür t RNA'nın kopyası vardır (toplam 200000 molekül). Amino asitler, tRNA'sına bağlanınca, antikodon da bazı değişiklikler olsa bile, kodona bağlanmada önemli bir aksama olmaz.
Sentetase enzimi (M.A. : 40 000-100 000) değişik yapılı bir moleküldür. Her biri on enzim içeren iki gruba ayrılırlar. Biri, t RNA'nın akseptör heliksinin minor oluğu ve diğeri de aynı heliksin büyük oluğu ile reaksiyon verir. Sentetase tarafından doğru amino asitin tanınması iki aşamada gerçekleştirilir.
1) Amino asit + ATP®amino acyl-AMP+PP1 ve
2) Amino acyl-AMP + RNA ® amino acyl t RNA + AMP
Sonuç: Amino asit + ATP + t RNA ® amino acyl t RNA + AMP + PP1
Sentetase enzimi, kendine ait amino asitle çok çabuk bağ kurar. Doğru olmayan amino acyl adenilatın oluştuğu veya yanlış amino asitin sentetase ile birleştiği durumlarda, sentetase enzimi geriye dönük düzeltme okuması (proof reading) yaparak, yanlış amino asiti hidrolize ederek çıkarır ve yerine doğrusunu koyar.
Kodon ve antikodon, genellikle 5' ® 3' yönünde okunur. Şöyle ki;
¾¾¾¾¾¾®
Kodon : 5' A C G -3' ise
Antikodon : 3' U G C -5'
¬¾¾¾¾¾¾
Bu durumda 1. baz (kodonda ) ile 3. baz (antikodon da) birbirinin partneri olurlar. Böylece, G ile C ve A ile U ile birleşirler. Ancak, G ile U da birleşebilir (Wooble birleşmesi). İkinci bazlar da aynı kurala uysalar bile bazen 3. pozisyonundaki bazlar değişik birleşme yapabilirler.

Transfer RNA'da birçok modifiye bazlar bulunur. Bu durum 4 bazın birinde oluşan modifikasyonlar sonu ortaya çıkar. Bazen böyle bazlar 50'ye ulaşabilir.
Antikodon bazlarında oluşan değişmeler, sekans değişikliklerine ve bunlar da okuma özelliğinin farklı olmasına yol açar. Hatta, modifikasyonlar antikodonun yakınında olsalar bile yinede baz birleşmeleri üzerine etkili olabilirler. Antikodonunun ilk pozisyonunda Inosine ekseriye bulunabilir 3 bazdan (U,C,A) biri ile birleşebilir.
Glycine tRNA ’sında bulunan antikodon bölgesindeki 3. baz I , Wooble nukleotid olup U,C,A ile bağlanabilirliği yandaki şekilde gösterilmektedir. | 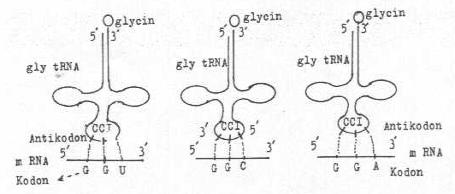
|
[1] Kaynak : Temel Mikrobiyoloji