Virusların Morfolojik ve Kimyasal Özellikleri
Prof. Dr. Mustafa Arda
Ankara Üniversitesi Veteriner Fakültesi
Temel Mikrobiyoloji 1
01. Virusların Morfolojik Özellikleri
01.01. Kapsomerler ve Kapsid
01.02. Zarf
01.03. Viral Nukleik Asitler (viral genom, DNA ve RNA)
02. Virusların Kimyasal Yapıları
02.01. Viral Proteinler
02.02. Lipidler
02.03. Karbonhidratlar
02.04. Fosfatlar
02.05. Nukleik Asitler (viral genom)
01. Virusların Morfolojik Özellikleri
İnsan ve hayvanlarda infeksiyon ve/veya hastalık oluşturan viruslar (genel bir terim olarak, hayvan virusları) morfolojik özellikleri yönünden fazla çeşitlilik göstermemekle beraber, elektron mikroskopik muayenelerde olgun viruslarda bazı farklı formlar gözlemlenmiştir.
Normal ışık mikroskopları ile görülebilen (1000 x veya 1500 x büyütmeli) Poxviridae familyası virusları hariç tutulursa, diğer virusların morfolojik karakterleri (olgun virus partiküllerinin genel görünümü ve yapıları) hakkında ayrıntılı bilgiler ve görüntüler, ancak, elektron mikroskopların keşfinden ve viroloji alanında kullanılmaya başlamasından sonra elde edilebilmiştir. Negatif boyama, X- ışınları difraksiyon, krioelektron mikroskopi, elektron mikroskop (TEM, SEM) ve diğer tekniklerin uygulanması, virusların daha net, ayrıntılı ve açık görüntülenmesine çok büyük katkısı olmuştur.
Yapılan çalışmalarla virusların başlıca 4 morfolojik form gösterdikleri belirlenmiştir.
1) Yuvarlak (sferik) formlar: İkosahedral simetriye sahip bazı DNA (Adenoviridae, Herpesviridae, Papovaviridae, Iridoviridae, Hepadnaviridae) ve RNA virus familyaları (Birnaviridae, Caliciviridae, Picornaviridae, Reoviridae ve diğer bazı familyalar) ile helikal simetriye sahip olanlar (Arenaviridae, Bunyaviridae, Coronaviridae, Orthoymxoviridae, vs) bu grup içinde yer almaktadırlar.
2) Flamentöz formlar: Filoviridae virusları (Ebola ve Marburg virusları) flamentöz morfolojik bir özellik gösterirler.
3) Mermi benzeri formlar: Rhabdoviridae virusları (kuduz virusu) mermi benzeri formlara sahiptirler.
4) Briket (tuğla) benzeri formlar: Poxviridae virusları (variola, vaccinia, Cowpox, Orf, BPS, Molluscum contagiosum, vs) bu gruba dahildirler.
Her ne kadar hayvan virusları başlıca 4 temel form gösteriyorlarsa da sferik olanlar daha fazla familya ile temsil edilmekte ve diğer 3 grup ise birer familyada bulunmaktadırlar.
Bakteri viruslarında (bakteriyofajlar): Bradley klasifikasyonunda, A,B, ve C grupları) rastlanan kuyruklu formlar, hayvan viruslarında görülmemektedir.
Yuvarlak DNA viruslarının boyutları 18-300 nm (nanometre) arasında değişmektedir (parvoviruslar, 18-26 nm ve iridoviruslar, 125-300 nm); RNA virusları, 25-300 nm (picornaviruslar, 25-30 nm ve paramyxoviruslar, 150-300 nm); Filoviruslar, 80 x 79-970 nm; rhabdoviruslar, 75 x 180 nm ve poxviruslar, 140-170 x 230-350 nm arası ölçülere sahiptirler.
Aynı familya içinde bulunan cinslere ait virusların boyutları arasında farklar bulunmaktadır.
Her ne kadar virusların bireysel formlarını ışık mikroskopları ile görüntülemek olası değilse de (poxvirusları hariç), ancak, bunların hücre içinde ürerlerken veya üredikten sonra oluşturdukları intrasitoplasmik veya intranukleer inklusiyon cisimciklerini görmek mümkündür. Işık mikroskopları ile kolayca gözlemlenen bu cisimcikler, ya sadece intrasitoplasmik (çiçek virusu, kuduz virusu, vs) veya sadece intranukleer (adenoviruslar, herpesviruslar, papovaviruslar, vs) oluşabilecekleri gibi her iki bölgede de aynı anda lokalizasyon gösterebilirler (CMV, kızamık virusu).
Hayvan virusları, elektron mikroskopla saptanabilen başlıca 3 temel yapısal karakter göstermektedirler.
1) Kapsomerler ve kapsid
2) Zarf
3) Nukleik asitler (viral genom, DNA ve RNA)
01.01. Kapsomerler ve Kapsid
Hayvan viruslarının genetik materyallerinin (DNA ve RNA) etraflarında belli sayıda ve birbirleri ile non kovalent bağlarla birleşmiş protein alt üniteleri (kapsomerler) bulunmaktadır. Kapsomerler, ikosahedral simetriye sahip viruslarda, belli bir düzen içinde yanana gelerek birleşir ve böylece genomun etrafında proteinden bir muhafaza oluşturur ki buna kapsid adı verilir. Helikal simetrili viruslarda ise, kapsomerler, viral nukleik asitin üzerinde yan yana gelmiş ve genoma bağlanmış durumdadırlar.
Viruslar, kapsid simetrilerine göre başlıca iki kısma ayrılmaktadırlar. Ancak, bu iki temel gruba uymayan poxvirusları 3. bir bölüm içinde toplanmış ve böylece hayvan virusları 3 kısımda incelenmektedirler.
1) İkosahedral (kübik) simetri
2) Helikal (sarmal) simetri
3) Kompleks yapı
1)İkosahedral (kübik) simetri: Kapsomerlerin yan yana gelerek oluşturdukları düzenli formlar arasında kübik simetrinin özel bir önemi bulunmaktadır. İkosahedral simetri gösteren bir kapsid (isometrik kapsid), 12 köşe, 20 eşkenar üçgen yüzey ve 30 kenardan oluşmaktadır. Kapsidin köşelerinde yer alan her bir kapsomer 5 tane komşu kapsomerle (pentamer, penton), kenar ile yüzeylerde bulunan her bir kapsomerde 6 tane komşu kapsomerle (hekzamer, hekzon) çevrilmiştir. Bunlar bazı viruslarda aynı ve bir kısımlarında ise farklı yapıda polipeptidlerden oluşmaktadır. Adenoviruslarında da köşelerden çıkan, kısa ve uçları şişkince (tokmak benzeri) uzantılar (fiber) bulunmaktadır. Protein karakterinde olan fiberler, virusun hücrelere tutunmasında görev alırlar.
Viruslara şekil veren, koruyan, viral genomun hücre içine girmesine yardımcı olan kapsid, protein yapısında olması nedeniyle de çok iyi bir antijeniteye sahiptir.
Kapsidlerin eşkenar üçgenlerinin kenarlarında eşit sayıda kapsomer bulunur. Üçgenlerin her biri de kendi içinde daha küçük üçgenlere ayrılabilir üçgenleşme (triangulasyon). Eşkenar üçgenlerin bir kenarındaki kapsomer sayısı (n) bilinirse, kapsiddeki toplam kapsomer sayısı (N) hesaplanabilir. Örn, adenoviruslarında her bir kenarda 6 kapsomer bulunur. Buna göre kapsidin toplam kapsomer sayısı aşağıdaki formül yardımı ile saptanabilir.
N = 10 (n-1)2 + 2
N = 10 (6-1)2 + 2 = 252
Kübik simetrili viruslarda nukleik asitler, kapsidin orta kısmında lokalize olmuştur. Genom ile kapsidin oluşturduğu birliğe, genellikle, nukleokapsid adı verilmektedir.
Bazı DNA ve RNA viruslarının morfolojik özellikleri aşağıdaki tabloda gösterilmiştir.
DNA Virusları (familyalar) | Çap (nm) |
Zarf
|
Simetri
| Kapsomer | Etere duyarsızlık |
Adenoviridae Hepadnaviridae Herpesviridae İridoviridae Papovaviridae Parvoviridae Poxviridae | 70-90 42 150 125-300 45-55 18-26 230x400 | - + + + - - + | İkosahedral İkosahedral İkosahedral İkosahedral İkosahedral İkosahedral Kompleks yapı | 252 ? 162 1892 72 32 ? | Duyarsız Duyarlı Duyarlı Duyarlı Duyarsız Duyarsız Duyarlı ve bazı türler duyarsız |
RNA Virusları (familyalar) |
|---|
Arenaviridae Birnaviridae Bunyaviridae Calicivirade Coronaviridae Filoviridae Flaviviridae Orthomyxoviridae Paramyxoviridae Picornaviridae Reoviridae Retroviridae Rhabdoviridae Togaviridae | 110-130 60 90-120 35-40 75-160 790-970x80 40-50 80-120 150-300 25-30 60-80 80-100 75x180 50-70 | + - + - + + + + + - - + + + | Helikal İkosahedral Helikal İkosahedral Helikal Helikal İkosahedral Helikal Helikal İkosahedral İkosahedral Helikal Helikal İkosahedral | ? 92 ? 32 ? ? ? ? ? 32 32,92 ? ? 60 | Duyarlı Duyarsız Duyarlı Duyarsız Duyarlı Duyarlı Duyarlı Duyarlı Duyarlı Duyarsız Duyarsız Duyarlı Duyarlı Duyarlı |
İkosahedral simetrili viruslara, hem DNA ve hem de RNA virus familyalarının bazılarında rastlanmaktadır. DNA viruslarından herpes ve hepadnavirusları, ile RNA viruslarından togavirus ve flaviviruslarının etrafında (kapsidin dışında) diğer bir muhafaza daha bulur(zarf).
Bazı virusların morfolojik ve genom özellikleri yandaki şekilde gösterilmektedir. | 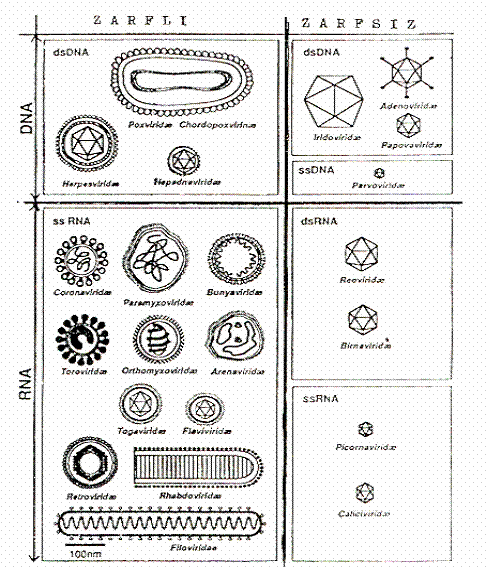 |
Virusların protein yapısındaki kapsidlerinde, farklı familyalara ait olanlarınki ile kros reaksiyon vermeyen ve vücutta spesifik immunolojik yanıtı stimule eden tip spesifik protein molekülleri vardır. Bunlar, SDS-PAGE'de (sodium dodecyl sulfate - polyacrylamide gel electrophoresis) molekül ağırlıklarına göre yapılan separasyonlarda jel üzerinde farklı aralıklarla oluşan bant düzenleri virus familya ve cinslerini belirlemede önemli rol oynarlar. Ayrıca, penton ve hekzonlarda veya aynı kapsid üzerinde değişik yerlerinde lokalize olmuş spesifik antijenik protein molekülleri bulunabilir ve bunlar teşhiste önemli fonksiyonlara sahiptirler. Adenoviruslarının fiberleri de aynı şekilde spesifik antijenik uyarım yapabilecek güçtedirler.
Reovirusların kapsidin etrafında iki adet konsantrik ve proteinden yapılmış muhafaza bulunmaktadır. İki muhafazalı olan virion infeksiyöz bir özellik taşır.
2) Helikal (sarmal) simetri: Helikal simetrili viruslara RNA virus familyalarında rastlanmaktadır. DNA viruslarında bu form bulunmamaktadır.
Sarmal simetrili viruslarda spiral formda bulunan genetik materyalin üzerinde, bir tür polipeptidden oluşan, oval veya yuvarlak şekilli kapsomerler yan yana gelerek genomla birleşmişlerdir. Böylece, nukleik asit (RNA) kapsomerlerle birlikte helikal (spiral) bir form alır (helikal simetri). Kapsomerler, RNA genomla birlikte olduklarından da helikal nukleokapsid özelliği taşır. Helikal simetri gösteren hayvan viruslarının nukleokapsidlerinin etraflarında ayrıca sellüler lipidlerden zengin bir zarf bulunur (zarflı viruslar). Nukleokapsid bazı viruslarda bu zarf içinde ikincil kıvrımlar (yumak gibi) yapabilir. Hayvan viruslarının aksine, bitki viruslarından olan, tütün mozaik virusu (TMV) zarf taşımaz (zarfsızdır). TMV, sert ve nukleokapsid uzun (çomak gibi), düz ve RNA etrafında, her biri tek bir polipeptidden oluşan, yaklaşık 2130 oval kapsomer bulunur.
3) Kompleks yapı formu: Yukarda açıklanan temel iki kapsid simetrisine uymayan Poxviridae familyası virusları kompleks yapı formu grubu içinde incelenmektedirler. Böyle yapıya sahip viruslarının yüzeylerinde tübüler elementler bulunur. Çiçek viruslarında, DNA karakterindeki viral genom ortada ve bikonkav bir lokalizasyon gösterir. Ayrıca, orta kısımlarda da lateral cisimcikler yer almışlardır (orthopoxvirusları). Parapoxviruslarının yüzeylerinde çapraz strüktürler bulunur. Poxviruslarının etraflarında lipidden zengin zarf vardır.
Virusların başlıca yapısal formları yandaki şekilde gösterilmektedir. | 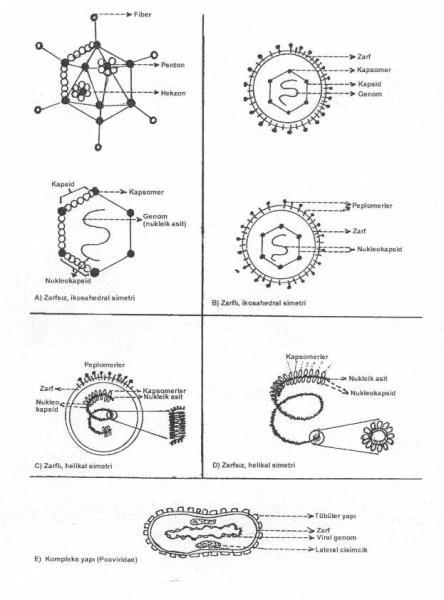
|
01.02. Zarf
Bazı DNA (herpesvirusları, hepadnaviruslar, poxvirusları) ve RNA virusları (arenavirusları, bunyavirusları ve diğerleri) nukleokapsidlerinin etraflarında, viruslar hücrelerden tomurcuklanarak olgunlaştıkları sırada hücreye ait membranlara sarılarak dışarı salınırlar. Bu nedenle de zarfın yapısı, hücre membranlarının (sitoplasmik membran, nükleer membran, endoplasmik retikulum) kimyasal yapısı ile çok büyük benzerlik gösterir. İki katmanlı (bilayer) olan zarf, hücreler tarafından kodlanmasına karşın, zarfta bulunan peplomerler (glikoprotein yapısında, spike) ise viruslar tarafından spesifiye edilirler. Tomurcuklanarak hücrelerden çıkan zarflı viruslar hücrelere zarar vermezler ve hücreler normal yaşamlarını sürdürürler. Diğer bir ifade ile, hücrelerde morfolojik değişiklikler (sitopatik efektler, CPE) yapmazlar. Ayrıca, böyle viruslar, persistent infeksiyonlara yol açabilirler. Buna karşın hücre içinde olgunlaşan bazı viruslar da hücrelerde parçalanma (sitolizis) yaparak dışarı çıkarlar.
Bilayer zarfın yüzeyinde lokalize olan ve viruslarca kodlanan peplomerler (spike; hemaglutinin, neuraminidaz, F-proteini), protein ve glikoprotein yapısında olduklarından da iyi bir antijeniteye sahiptirler. Vücutta, spesifik immunolojik bir yanıt oluştururlar. Peplomerler, elektron mikrografide çıkıntılar halinde görülürler. Bunlardan ayrı olarak ta bazı virusların (rhabdoviruslar, reoviruslar, orthomyxoviruslar, vs) zarflarının iç yüzeylerinde ve zarfın sağlamlığını destekleyen nonglikozile matriks proteini (M proteini) yer almaktadır.
Viruslarda kapsid ve zarfın çok önemli fonksiyonları bulunmaktadır. Bunlardan bazıları aşağıda belirtilmiştir,
a) Kapsidler ve zarflar, virusları, hem hücre içi (nukleazlardan) ve vücut içi antiviral maddelerden ve hem de vücut dışı fiziksel, kimyasal ve diğer virusidal faktörlerin zararlı etkilerinden de korurlar,
b) Kapsidler ve zarflar (peplomerler), protein ve glikoprotein yapısında olduklarından ve virus grupları arasında farklı kimyasal özellik taşıdıklarından, vücutta spesifik bağışıklığı uyararak özgül antikor sentezini sağlarlar,
c) Kapsidler ve zarflar, hücre yüzeylerindeki spesifik reseptörlere bağlanmada önemli rol oynarlar ve virusların infeksiyon oluşturmasının veya hücrelere girişinin de ilk basamağını sağlarlar,
d) Kapsidler ve zarflar, virusların güvenli olarak sitoplasmik membranı geçmesini ve hücre içine (sitoplasmaya) ulaşmasına yardımcı olurlar,
e) Kapsidler ve zarflar, virusların morfolojik özelliklerini belirlemede de fonksiyoneldirler,
f) Kapsidler ve zarflar (peplomerler), vücutta oluşan spesifik antikorların tanınmasında ve bunlara bağlanmada ve antikor-antijen interaksiyonlarında önemli görevlere sahiptirler.
g) Bazı poxvirusları hariç tutulursa, zarfın bütünlüğü infektivite için gereklidir.
Zarflı viruslarda, glikoprotein yapısında olan peplomerlerin yanı sıra Transport kanal proteinlerinin varlığı da bildirilmektedir. Bunlar, membranın (zarfın) permeabilitesinde değişiklikler yapmakta, virionun internal ortamını modifiye etmekte ve virionun olgunlaşmasına katkıda bulunmaktadırlar.
01.03. Viral Nukleik Asitler (viral genom, DNA ve RNA)
Virusların genetik yapılarını oluşturan nukleik asitler (viral genom), deoksiribonukleik asit(DNA) veya ribonukleik asit (RNA)lerden sadece birinden oluşur. Diğer bir ifade ile, virionda ya DNA veya RNA'lardan sadece biri vardır ikisi birden bulunmamaktadır. Bunlara karşın, bakterilerin genetik materyallerini hem DNA ve hem de RNA oluşturur. Ancak bunlardan mRNA'da genetik bilgiler bulunur. Diğer RNA'lar (tRNA ve rRNA) nongenomiktirler.
Viral genom, hücre içinde kendi replikasyonunda fonksiyonu olan enzimlerden bazılarının kodlarına sahip olmasına karşın, gerekli diğer enzimleri, makromolekülleri, replikasyon ve ekspresyon mekanizmalarını, içinde üredikleri hücrelerden sağlarlar ve yararlanırlar.
Bazı nukleik asitler, hücrelere girdiklerinde, infeksiyonu başlatabilir ve infeksiyöz yeni nesiller oluşturabilir. Buna karşın, bir kısım viruslar da, genetik informasyonların hemen hepsi bulunduğu halde, viral genom infeksiyöz değildir. RNA viruslarında, viral genom, eğer negatif (sens) polariteli ise, veya çift iplikçikli ise, bunlardan pozitif polariteli mRNA( +mRNA)'nın meydana gelebilmesi (transkripsiyonu) için viriona bağımlı transkriptaz enzimine gereksinim vardır. Bazı viruslarda da, transkripsiyonda sellüler transkriptaz kullanılır. Retrovirusların pozitif polariteli iki molekül tek iplikçik RNA'ları infeksiyöz bir özellikte değildir. Bu virusların ancak, proviral çift iplikçikli DNA'ları hücre genomuna integre olduktan sonra replikasyonları başlar. Retroviruslar hariç, diğer virusların genomlarında, her genin sadece bir kopyası bulunur.
Viral genom, taşıdıkları genetik materyal türü yönünden başlıca iki kısma ayrılırlar.
1) Deoksiribonukleik asit (DNA): Bütün DNA viruslarında tek bir molekül genom bulunur. Bu da ya tek iplikçikli (ssDNA, parvoviruslar) veya çift iplikçikli (dsDNA), lineer veya sirküler bir özelliğe sahiptirler. Bunlar hakkında ayrıntılı bilgiler tablo 44.2 gösterilmiştir. Papovaviruslarında sirküler olan dsDNA, genellikle, süpersarmal (süperheliks) bir form gösterir. Çift iplikçikli ve sirküler hepadnaviruslarında DNA iplikçiklerinden biri diğerinden daha kısadır ve bunu bağlı olarak ta genom kısmen tek iplikçikli bir özellik taşır (ss/dsDNA). Poxvirusların lineer iki iplikçiğinin uçları birbirlerine kovalent bağlıdır ve denatürasyon durumunda sirküler tek iplikçik form oluşmaktadır. Lineer çift iplikçikli bazı herpesviruslarında genomun sonunda tekrarlanan sekanslar vardır. Adenoviruslarında, terminal tekrarlar birbirlerine ters oriyentasyon gösterirler (inverted terminal repeat, ITR). Parvoviruslarında da uçlarda palindromik sekanslar bulunur ve bunlar bir saç tokası görünümü alabilmektedirler. Terminal strüktürler yapışkan uçlar oluşturmaları yönünden de önemlidir.
DNA viruslarında, bazı RNA viruslarında rastlanan segment oluşumu, saptanamamıştır. Bazı DNA virusları, kendi nukleik asitlerin sentezinde önemli rolleri olan proteinlerin (özellikle, enzimlerin) kodlarını taşırlar. Örn, hepadnaviruslar DNA'ya bağımlı DNA polimeraz ve poxvirusları da çok sayıda (polimeraz, nukleertopoizomeraz, fosfohidrolaz, vs gibi enzimlerin kodlarına sahiptir.
DNA viruslarında genom büyüklüğü, küçük DNA viruslarında (parvoviruslar) 4-5 kb (MA: 1.5-2.2 x 106) ve büyük DNA viruslarında (poxvirusları) 130-280 kbp (MA: 130-200 x 106) arası bir varyasyon gösterir. Genellikle, 1 kb veya 1 kbp uzunluk, ortalama büyüklükte bir proteini kodlayacak bilgilere sahip olduğu kabul edilir. Bu nedenle de genomları 4-200 kb uzunlukta olanlar hem 4-200 gene ve dolayısıyla da 4-200 proteini kodlayabilecek kapasiteye sahip olmaktadırlar. Ancak, kodlamayan sekanslar (intron), tekrarlanan baz sıraları ve diğer nedenler göz önüne alınırsa, yukarıdaki görüş pek gerçekleşememektedir. Poxvirusları hariç tutulursa, DNA viruslarının hemen hepsi ikosahedral simetriye sahiptirler.
Bazı DNA ve RNAViruslarının Genom Karakterleri
DNA Virusları (familyalar) | Mol. Ağ. (x106) | Büyüklüğü (kb, kbp) | Nükleik asid özellikleri | Gen sayısı (yaklaş.) | Segment sayısı | Enzim |
Adenoviridae Hepadnaviridae Herpesviridae İridoviridae Papovaviridae Parvoviridae Poxviridae | 20-30 1.6 90-130 150 3-5 1.5-2.2 130-200 | 30-37 3.2 120-220 150 5.8 5 130-280 | ds lineer ds sirküler ds lineer ds lineer ds sirküler ss(-) lineer ds lineer | 30 4 160 5-8 3-4 300 | 1 1 1 1 1 1 1 | - +(RT) - + - - + |
RNA Virusları (familyalar) |
Arenaviridae Birnaviridae Bunyaviridae Caliciviridae Coronaviridae Filoviridae Flaviviridae Orthomyxoviridae Paramyxoviridae Picornaviridae Reoviridae Retroviridae Rhabdoviridae Togaviridae | 3-5 6-15 2.6 7 13 4 5 5-7 2.3-2.8 12-15 7-10 4 4 | 10-14 7 13.5-21 8 27-33 12.7 10 13.6 18-20 7.5-8.5 18-27 7-10 13-16 12 | ss(-) lineer ds lineer ss(-) lineer ss(+) lineer ss(+) lineer ss(-) lineer ss(+) lineer ss(-) lineer ss(-) lineer ss(+) lineer ds lineer ss(+) lineer ss(-) lineer ss(+) lineer | 10 3 4-6 30 10 10 10 4-6 10-12 4 5 5 | 2 2 3 1 1 1 1 7,8 1 1 10,11,12 2 1 1 | + + + - - + - + + + + + (RT) + - |
ds : Çift iplikçik Jawetz, 1998;Ustaçelebi, 1993;Fenner, 1993, Akan, 1994
ss : Tek iplikçik
+ : Pozitif polarite (+ sens)
- : Negatif polarite (- sens)
RT: Reverse transkriptaz
2) Ribonukleik asit (RNA): RNA karakterinde genoma sahip viruslar, nukleik asitlerinin yapısal özellikleri yönünden, DNA viruslarından daha komplike bir durum gösterirler. Bunlar da, tek veya çift iplikçikli, lineer, segmentli ve segmentsiz bir yapısal organizasyona sahiptirler. RNA viruslarında sirküler genom bulunmamaktadır: Retroviruslarında genom, iki adet tek iplikçik ve pozitif polariteli RNA molekül yapısındadır. Bazı virus familyalarındaki (Bunyaviridae, Orthomyxoviridae, Reoviridae, vs) viruslarda bulunan segmentlerin her biri ayrı bir gen olduğu ve segmentlerin birbirlerinden tam ayrı olmadığı ve birbirlerine non kovalent bağlarla birleştiği kabul edilmektedir.
RNA viruslarında genom hem ikosahedral ve hem de helikal simetri gösterir. Tek iplikçik RNA viruslarından picornaviruslar ve caliciviruslar dışındaki, ssRNA taşıyan virusların etraflarında lipidden zengin bilayer bir zarf bulunur.
ssRNA virusları, viral nukleik asitlerinin polarite durumlarına göre pozitif (+) veya negatif (-) polarite (sens) olmak üzere iki kategoriye ayrılmaktadırlar. Eğer RNA hücre içinde mRNA fonksiyonuna sahipse, pozitif polariteli (+) RNA ve eğer böyle bir etkinliği bulunmuyorsa negatif polariteli olarak (-) RNA olarak kabul edilir.
Viral RNA'nın (+) polariteli olduğu durumlarda, genomun 3'-ucu, genellikle, poliadenilatlı (pol AAA....) bulunmaktadır (örn, caliciviruslar, coronaviruslar, picornaviruslar, togaviruslar); bazılarının 5'-ucu ise kepli (cap)'dir (7-metil guanosin). Örn, coronaviruslar, flaviviruslar, togaviruslar, vs). Picornavirusları ve caliciviruslarda, RNA'nın 5'-terminusunda protein molekülü vardır.
RNA viruslarında replikasyonda görev alan bazı enzimlerin kodları yer almaktadır. Örn, retroviruslarında, RNA'ya bağımlı DNA polimeraz; orthomyxovirusları, paramyxo-filo, bunya, reo ve arenaviruslarında, RNA'ya bağımlı RNA polimeraz enzimi gibi.
RNA viruslarının genom büyüklüğü, 7,5 kb (picornavirusları, MA: 2.3-2.8 x 106) ile 27-33 kb (coronaviruslar, 7 x 106) arasında değişmektedir.
DNA ve RNA viruslarında genoma ait bazı özellikler ile viral genom türleri yandaki şekilde gösterilmektedir.
| 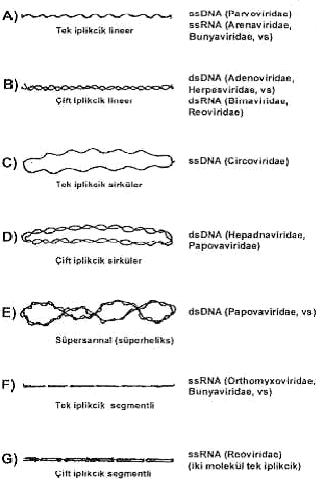 |
02. Virusların Kimyasal Yapıları
Olgun bir virus partikülü, her ne kadar bir bakteri kadar zengin ve çeşitli kimyasal komponentlere sahip değilse de duyarlı organizmaya girdiklerinde değişik derecede (virusa ve konakçıya bağlı olarak) immunolojik bir yanıt oluşturabilecek moleküllere ve kompleks yapılara sahip bulunmaktadırlar. Bu nedenle de, viruslar, immunolojik özellikleri bakımından, bakterilerden hiç de gerilerde değildir ve hatta daha da önde oldukları söylenebilir.
Virusların kimyasal yapılarını oluşturan başlıca komponentler hakkında aşağıda kısa ve özlü bilgiler verilmektedir.
02.01. Viral Proteinler
Virusların yapılarında bulunan proteinler (viral proteinler) başlıca iki karakter taşımaktadırlar.
1) Yapısal (strüktürel) proteinler: Bu tür proteinler virionun bir çok bölgesinde lokalize olmuşlardır. Bunlardan, kapsid proteinleri, bazı DNA ve RNA viruslarında kapsidi oluşturan protein alt ünitelerinin (kapsomerler) yapısında bulunurlar ve çok iyi immunojenik aktiviteye sahiptirler. Kapsid proteinleri, bir virionun yüzeyinde, değişik karakterlerde ve lokalizasyonlarda olabilirler (VP1, VP2, VP3......,gp,....). Helikal simetrili viruslarda ise, genomla birleşik olan kapsomerler, nukleokapsidi oluştururlar. Burada da kapsomerlerin yapısı yine proteinden meydana geldiğinden antijenik bir özellik taşır.
Yapısal proteinlerin bazıları da virusun bağlanma proteinlerini (ligand) oluştururlar.
Zarflı viruslarda zarfın yapısında bulunan proteinler (zarf proteinleri) virus tarafından kodlanan peplomerlerde lokalize olmuşlardır. Peplomerlerde virus tarafından kodlanan peplomerlerde lokalize olmuşlardır. Peplomerlerde (spike), başlıca hemaglutinin, neuraminidase ve F-proteinleri bulunmakta ve bunlar, hücre yüzeylerinde yerleşik olan spesifik reseptörlerle bağlanabilen ligandları oluştururlar. Protein ve glikoprotein özelliği taşıyan bu moleküller de çok iyi antijenik karaktere sahiptirler. Hemaglutininler (trimer molekül) genellikle, hem alyuvarlara bağlanarak hemaglutinasyon fenomenine neden olurlar ve hem de hücrelere bağlanmada etkin ligandları oluştururlar. Neuraminidase (tetramer molekül) ise, hücre yüzeylerinde lokalize olan reseptörlerin yapısındaki oligosakkaridlerdeki sialik asidi hidrolize ederek bunun aktivitesine mani olur ve virusun bağlantısını çözerek serbest kalmasını sağlar (elüsyon). Bu nedenle neuraminidaz "reseptör parçalayan enzim" olarak tanınmaktadır (parapoxvirusları, orthopoxviruslar ve diğerleri). Virusların hücre içlerine girmesinde bu olgunun önemi fazladır. F-proteini (füsyon proteini), bu moleküller, hücrelerin bir araya gelmesini sağladığı gibi, hücre membranları ile de interaksiyona girerek zarfın membranla bütünleşmesinde de önemli rol oynar ve nukleokapsidin sitoplasmaya girişini kolaylaştırır. Zarflı olmasına karşın, Coronaviridae ve Herpesviridae viruslarındaki peplomerlerin etkinliği, Orthomyxoviridae ve Paramyxoviridae virusları kadar değildir ve oldukça zayıftır.
Bazı zarflı viruslarda da (rhabdoviruslar, reoviruslar, orthomyxoviruslar, vs) yapısal bir karakter taşıyan ve zarfın iç yüzünde lokalize olan nonglikozile matriks proteinleri (M-proteinleri) bulunmaktadır. Viruslara şekil vermede rollere sahip olan M-proteinlerine Arenaviridae, Bunyaviridae ve Coronaviridae familyası virusların da rastlanmamıştır.
Zarfın yapısı yandaki şekilde gösterilmektedir. | 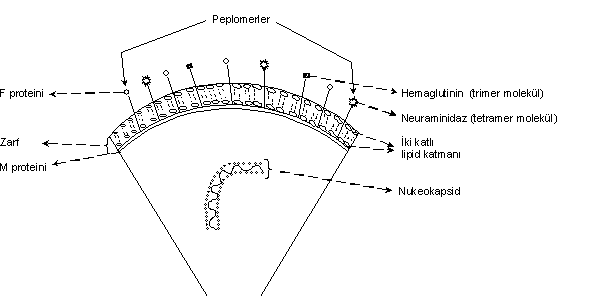
|
2) Yapısal olmayan (non strüktürel) proteinler: Bu proteinler, genellikle, viruslar tarafından kodlanmakta olup enzim karakteri göstermektedirler. Bunlar, virusların hücre içinde replikasyonları ve transkripsiyon regulasyonunda görev almaktadırlar (transkriptaz, revers transkriptaz, erken gen proteinleri ve diğerleri). Poxviridae familyası virusları kodladığı proteinleri bakımından oldukça zengindirler. Retroviruslar da revers transkriptaz enziminin kodlarına sahiptirler.
02.02. Lipidler
Bazı DNA (Herpesviridae, Hepadnaviridae, Iridoviridae) ve RNA virus familyalarında (Arenaviridae, Bunyaviridae ve diğerleri) nukleokapsidlerin dışında yer alan ve hücre orijinli olan zarflar genellikle lipidlerden zengindirler. Çünkü, zarflar, viruslar, hücre membranlarından (sitoplasmik membran, nükleer membran, endoplasmik retikulum) çıkarlarken (olgunlaşma sırasında) alırlar. Bu nedenle de zarfın kimyasal yapısı ile hücrelerinki arasında çok yakın bir benzerlik bulunmaktadır ve bunlar hücre tarafından kodlanırlar. Zarflarda bulunan lipidler, viruslara göre değişmek üzere, hücre kuru ağırlığının %20-35'i kadar olabilmektedirler. Herpes viruslarının zarfları, hücrelerin nükleer membranları, orthomyxovirus ve paramyxovirusların zarf lipidleri de sitoplasmik membranların kimyasal yapıları ile özdeştirler. Lipidlerin yapısında fosfolipid, kolesterol, nötral yağlar, trigliseridler, glikolipidler, vs. bulunmaktadır.
Virusların etrafında bulunan zarfların lipid kompozisyonları ve moleküler yapıları, virus türlerine ve ayrıca, içinde üredikleri hücreler (özellikle, olgunlaşarak çıktıkları membranların yapısına) göre değişiklik göstermektedir. Hatta, aynı tür virusun zarf lipid kompozisyonu, üredikleri hücreler farklı ise, değişik yapısal karakter gösterebilir.
Zarflar, genellikle iki katmanlıdır (bilayer).
02.03. Karbonhidratlar
Karbonhidratlar, virusların değişik bölgelerinde lokalize olmuşlardır. Virusların genetik materyallerinde (DNA ve RNA) pentoz şekerleri bulunmaktadır. DNA viruslarında 2-deoxy D-ribose olan şeker, RNA viruslarında D-ribose molekülü halindedir. Bu durumları ile de, genoma ad verirler (DNA ve RNA gibi).
Karbonhidratlar, genomun yapısındaki bazlar ile fosfat molekülleri arasında bağ kurarlar. Şöyle ki, pirimidin bazlarında, şekerin 1 no’lu karbon atomu ile bazın 3 no’lu nitrojeni ve pürin bazlarında da şekerin 1 no’lu karbon atomu ile, bazın 9 no’lu nitrojeni arasında beta glikozid bağları ile ve diğer taraftan da pentoz şekerleri 3. ve 5 no’lu karbon atomları ile fosfat moleküllerine fosfodiester bağları ile birleşerek kompleksler oluştururlar.
Zarfların yüzeyinde bulunan peplomerlerin yapısında da şeker molekülleri bulunduğundan bunlar, glikoprotein yapısı taşırlar ve iyi bir antijeniteye de sahiptirler. Buna karşın, matriks proteininde (M-proteini) şeker molekülü bulunmamaktadır (nonglikozile).
02.04. Fosfatlar
Fosfat molekülü (H3PO4), viral nukleik asitlerin 3. önemli yapısal komponentini oluşturur. Ayrıca, Poxviridae familyası viruslarında fosforilize olmuş bazı proteinlere (fosfoprotein) veya lipidlere (fosfolipid) rastlanılmaktadır.
02.05. Nukleik Asitler (viral genom)
Viruslar, genetik materyal olarak DNA (deoksiribonukleik asit) veya RNA (ribonukleik asit)'lardan sadece birini taşırlar. Bütün viral genomlarda her genin sadece bir kopyası olmasına karşın, retroviruslarında iki molekül tek iplikçik RNA bulunmaktadır (2, ss RNA, diploid).
Nukleik asitler, DNA veya RNA, 3 tür komponentten oluşmaktadır. Bunlar da,
1) Pirimidin veya pürin bazları: Pirimidin bazları timin (T), sitozin (C) ve pürin bazları adenin (A) ve guanin (G)'dir. Ancak, RNA'da timin yoktur bunun yerini urasil (U) almıştır.
2) Pentoz şekeri (C5H10O5) : DNA'nın yapısında 2-deoxy D-ribose (C5H10O4) bulunmasına karşın RNA'da D-ribose (C5H10O5) molekülü vardır.
3) Fosfat molekülü (H3PO4): DNA ve RNA'da aynı yapıda fosfat molekülü bulunmaktadır.
[1] Kaynak :Temel Mikrobiyoloji