Prof. Dr. Mustafa Arda
Ankara Üniversitesi Veteriner Fakültesi
Temel Mikrobiyoloji1
01. Genel Bilgiler
02. Sıvı Ortamda Üreme
03. Katı Ortamlarda Üreme
04. Üremenin Ölçülmesi
01. Genel Bilgiler
Mikroorganizmalar, uygun besi yeri ve çevresel koşullar altında, türlerine özgü bir süratle ürerler. Koşulların uygunluğu devam ettiği sürece, buna paralel olarak çoğalma da sürekli olur. Ancak, laboratuvarlarda, mikroorganizmaları üretmede sınırlı miktarda besi yerleri kullanıldığından, mikropların üremeleri kısıtlanır. Mikroplar üredikçe, ortamdaki gıda maddeleri azalır ve tükenir. Optimal koşulların değişmesi (pH, osmotik basınç, oksijen, yüzey gerilimi, vs.) ve besi yerinde toksik metabolitlerin birikmesi, miktarı az olan besi yerinde üremeyi kısa bir süre sonra baskılar ve durdurur. Örn, her 20 dakikada bir defa bölünerek üreyen E. coli hücresinden, uygun koşullar devam ettiği sürece 48 saat içinde 2114 (veya 2.2x1043) hücre meydana gelir.
Yukarıda bildirilen nedenlerle üremenin kısıtlanacağı, aynı zamanda her hücrenin tekrar bölünme olasılığının sınırlanacağını ve her mikrop için bölünme süresinin 20 dakika olmayacağını düşünülürse, bildirilen rakamlara ulaşmanın güçlüğü ortaya çıkar.
İnsan ve hayvanlarda hastalık oluşturan yuvarlak (streptokok, stafilokok, diplokok, Neisseriae 'lar, vs.), çomak bakteriler (pastörella, korinebakteri, mikobakteri, salmonella, brusella, vibrio, vs.), aerobik basiller (B. anthracis, B. subtilis, B. cereus, vs.), klostridium sınıfına ait bakteriler (C. tetani, C. botulinum, C. septicum, C. welchii, vs.) ve sarmal biçimdeki (spiroketler) mikroplar kısa eksen boyunca ortadan ikiye bölünmek suretiyle ürerler. Sporlu olan aerobik ve anaerobik mikroplarda sporlar üreme aracı değildirler. Bu basillerin vegetatif formları da aynı şekilde ortadan ikiye bölünmek suretiyle çoğalırlar. | 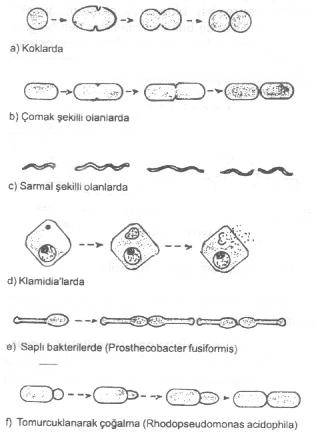
|
Bölünme başlamadan önce, bakteri, iki kardeş hücreye yetecek kadar enzimleri, gerekli diğer organik ve inorganik maddeleri hazırlar ve biriktirir. Bu işlemler yapılırken hücre içinde özellikle nukleer bölgede bir organizasyon görülür. Toplu halde bulunan nukleus orta bölgede uzamağa başlar. Nukleus sitoplasmik membrandaki özel yere (muhtemelen mesosom) bağlanarak replikasyona başlar. Replikasyon tamamlanınca, hücre duvarından içeri doğru ve karşılıklı olarak bir septum oluşumu görülür.
Buna, sitoplasmik membran da iştirak eder ve septumlar içeri doğru uzayarak hücreyi ortasından iki kardeş hücreye ayırır. Bu iki hücre birbirinden ayrılarak tam bağımsız hale gelirler veya birbirlerine bitişik olarak kalırlar (streptokok, streptobasil, flamentöz formlar. | 
|
Watson Crick modeli DNA yapısı (çift iplikçikli ve sarmal, dupleks) replikasyonun, semi-konservatif bir şekilde olabileceğini ifade eder. Bu görüşe göre, her bir iplikçik, kendine komplementer olan diğer yeni iplikçiğin sentezlenmesinde, kalıp ödevini görür. Replikasyon genellikle, sabit ve belli bir yerden başlayarak diğer uca doğru gider. İplikçikler birbirinden ayrılırken, karşılarında, birbirini tamamlayan ve homolog olan yeni iplikçik sentezlenmeye başlar ve bu işlem, tüm iplik ayrılıncaya dek devam eder. Böylece DNA replike olur ve oluşan çift iplikçiklerden biri (ki bunda parental DNA'dan da bir iplikçik vardır) kardeş hücreye geçer, diğeri ise parental hücrede kalır. Bu nedenle F2 generasyonda, çift iplikçikte bir tane orijinal ve bir tanede yeni sentezlenen iplikçik bulunur.
E. coli 'de kromozom sirküler şekildedir. Tam bir duplikasyon için 20-30 dakika geçmekte ve heliks bu süre içinde açılmaktadır. DNA, her 3.4 nm uzunlukta bir büküme sahip olduğu düşünülürse ve bir kromozomun da, bakteri türüne göre değişmek üzere, ortalama 1 mm. uzunlukta olabileceği hesaba katılırsa, bir bölünme süresi (20-30 dakika) içinde, DNA'nın ne kadar hızla açılması gerekeceği ortaya çıkar. DNA bir taraftan açılırken diğer taraftan da sentezlenen yeni iplikçik nedeniyle tekrar heliks formunu almakta ve kapanmaktadır. Bu açılma işlemi, düz veya lineer yapıda olanlarda veya sirküler olmayan kromozomlarda çok daha basitçedir. Yapılan çalışmalarda, DNA iplikçiklerinin birinde başlatma yerinin (replikatör) bulunduğunu ve daima da buradan açılmaya başladığını ve sona doğru ulaştığını ortaya koymuştur. Replikasyonu başlatan ve heliksi açan özel proteinler de bulunmaktadır (helikase).
Genetik kontrol altında bulunan DNA replikasyonu bazı hallerde bozulabilir. DNA 'nın replikasyonu tamamlanınca septum oluşumunun gecikmesi veya replikasyon çatalının fazla olması hallerinde iki adet çekirdek bulunabilir. Ancak bu durum çok nadiren meydana gelir.
02. Sıvı Ortamda Üreme
Mikroorganizmalar optimal koşulları içeren sıvı ortamlarda, katı besi yerlerinden daha çabuk ürerler. Üremenin hızı, mikrop türüne özgü genetik bir karakter olmakla beraber, besi yerinin bileşimi ve çevresel koşullarla da yakından ilişkilidir. Mikroorganizmalar, ikiye bölünmek suretiyle geometrik bir üreme tarzı gösterirler.
1 | —› | 2 | —› | 4 | —› | 8 | —› | 16 | —› | 32 | —› | ... |
20 | — | 21 | —› | 22 | —› | 23 | —› | 24 | —› | 25 | —› | ... |
Bu geometrik üremenin matematiksel ifadesi de (1x2n)'dir. Bakteri populasyonunda meydana gelen her bölünmeye generasyon ve iki generasyon arasında geçen zamana da generasyon süresi adı verilir. Her generasyon sonunda populasyondaki fert sayısı iki misli artar. Generasyon süresi bakterilerde çok değişiklik gösterir. E. coli 'de 18-20 ve S. aureus 'da 27-30 dakika olan genarasyon süresi, Mycobacterium tuberculosis 'de 792-932 dakikadır.
Mikroorganizmalarda generasyon sayısını ve süresini hesaplamak mümkündür. Bunun için başlangıçta sıfır zamanda ekilen mikrop sayısını (a), aradan geçen zamanı (t) ve bu süre sonundaki mikrop sayısını (b) bilmek gereklidir. Optimal koşulları içeren bir besi yerine bir adet mikrop ekilirse belli bir süre (t) sonra mikrop sayısı (b) aşağıdaki tarzda bulunur;
b = a x 22 formülünde 2n, n kadar sayıdaki generasyon sonundaki bakteri sayısını ifade eder. Eğer sıfır zamanda, (a) sayıda mikrop ekilmişse, total mikrop sayısı (b),
b = a x 2n (veya Nn = 2n No) olur.Bu eşitlikten yararlanarak generasyon süresi (g) ve sayısı (n) hesaplanabilir. Yukarıdaki denklemin logaritması alınır.
log b = (log a + n. log 2) ; Buradan
n= (log b - log.a) / log 2 olur. Diğer taraftan
n = t / g olduğundan
(t / g) = (log b –log.a) / log 2 olur.
Bu eşitlikten yararlanılarak,
g = (t x log 2) / (log b –log a) elde edilir.
Örneğin, a = 103, b = 109 ve t = 10 saat ise,
n = 109 – 103 / 0,3 = 6 / 0,3 = 20 (generasyon sayısı)
g = 10 x 0,3 /109 – 103 = 3 / 6 = 1/2 saat (generasyon süresi)
elde edilen sonuçlara göre, her yarım saatte bir generasyon olmak üzere, 20 generasyon aradan geçmiş bulunmaktadır.
Generasyon süresi (g), spektrofotometrik yöntemlerle de saptanabilir. Buna göre yapılan hesaplamada, optik dansite (OD) esas alınır ve aynı formül uygulanır.
g = t x log.2 / ODb -ODa
Bu formülde, t süresi sonunda optikal densite (ODb) ile başlangıçtaki optikal densite (ODa) kullanılmıştır.
Uygun bir besiyerine ekilen bir mikroorganizmanın belli bir süre içindeki üreme durumu başlıca 4 evre gösterir. Yandaki şekilde a) Latent dönem, b) Üreme dönemi (logaritmik dönem), c) durma dönemi, d) ölme dönemi görülmektedir. | 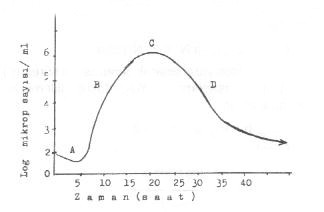
|
Bakterilerin Uygun Sıvı Ortamda Üremesi
A) Latent dönem: Bu fazda yeni veya değişik ortama ekilen mikroplarda üreme olmadığından sayıca da bir artma olmaz. Buna karşın, bakteriler yeni enzimler, metabolik intermedierler ve gerekli maddeleri sentezlediğinden, metabolizma artmıştır. Bu dönem, aynı zamanda, mikroorganizmaların, ortama adaptasyonlarını da sağlar. Adapte olamayanların ölmeleri nedeniyle mikrop sayısında azalmalar da görülebilir. Ortama uyabilen ve yeni maddeler hazırlayan mikroorganizmalar belli bir latent devreden sonra bölünerek üremeye başlarlar. Bu dönemin uzunluğu veya kısalığı mikropların türüne bağlı olmakla beraber, çevresel koşullara (pH, ısı, osmotik basınç, yüzey gerilimi, oksijen, vs.), besi yerinin bileşimine, ekilen mikropların ilk alındığı kültürünün dönemine de bağlıdır. İnokulum, bir önceki kültürün durma veya ölme döneminden alınmışsa, veya değişik bileşimdeki besi yerine ekilmişse veya aralarında ısı farkı varsa, bu takdirde uzun veya kısa bir latent dönem oluşabilir. Eğer her iki besi yeri aynı nitelikte ve ekilen mikroplar da bir önceki kültürün üreme döneminden alınmışsa, bu takdirde, latent devre oluşmaz ve mikroplar normal üremelerini sürdürürler.
Ortamda antimikrobial maddelerin bulunması veya hücre bölünmesi için gerekli maddelerin olmaması hallerinde bu faz çok uzun olur veya bölünme olmaz. Latent dönem, şekilde (A) harfi ile gösterilmiştir.
B) Üreme dönemi (logaritmik dönem): Yeterince ve gerekli maddeleri alan, sentezleyen ve latent fazı geçen mikroplar, kendi türlerine özgü bir generasyon süresi içinde ve belli aralıklarla bölünerek çoğalmaya başlarlar. Bakteri sayısı ve bölünme zamanla orantılı olarak artar. E. coli 'de bu süre 18-20 dakika arasında değişmesine karşın, mikobakterilerde 800-900 dakika kadardır. Ancak, aynı tür mikroorganizmalar, üreme döneminde olsalar bile, yine aralarında biraz geç veya biraz kısa süre içinde bölünenler bulunacaktır ve hatta hiçbir bölünmeyenler de olabileceği gibi. Bakteri hücresinin bölünmesi, biyokimyasal aktivite ile de paralellik gösterir.
Bu dönemdeki kültürler, belli zaman aralıklarında sayıma tabi tutulurlarsa üreme eğrisi düz veya dik bir durum gösterir (B). Bu fazda fizyolojik olarak çok aktif olan mikroorganizmalar, fiziksel ve kimyasal etkenlere karşı çok duyarlıdırlar. Mikroplar, bu dönemde, birçok karakterleri yönünden de bir örneklik gösterirler.
Üreme dönemi çok uzun bir süre devam etmez. Birkaç saat sonra bölünme hızında yavaşlama başlar. Ortamın sınırlı olması nedeniyle gıda ve enerji kaynaklarının sarf edilmesi ve azalması, metabolizma artıklarının ve toksik maddelerin birikmesi, ozmotik basıncın ve yüzey geriliminin değişmesi, oksijenin azalması, besi yerinde bulunan fermente olabilir karbonhidratların ayrışması sonu oluşan çeşitli organik asitlerin ortamın pH 'sını düşürmesi, vs. gibi başlıca faktörler olumsuz yönde etkileyerek üremenin yavaşlamasına sebep olurlar.
Mikroorganizmaları devamlı üreme döneminde tutabilmek için bazı özel aletler, kemostat ve türbidiostat, geliştirilmiş ve kullanılmıştır. Kemostat, kültüre, mikrobun bölünme durumuna göre, belli aralıklarla damla halinde veya muayyen miktarda taze besi yeri veren ve aynı miktar kullanılmış besi yerini dışarı salan bir alettir. Türbidiostat ise, fotoelektrik yöntemi ile kontrol edilen kültürlerde, üreme sonu oluşan bulanıklık belli bir düzeye ulaşınca, kendiliğinden ortama taze besi yeri veren ve aynı miktar kullanılmış vasatı dışarı çıkaran, bir aparattır.
C) Durma dönemi: Mikropların ürediği sınırlı miktardaki sıvı ortamdaki bozulan optimal koşullar değişmediği veya düzeltilmediği takdirde, üreme giderek yavaşlar. Bakteri populasyonu bir süre sayıca değişmeden kalır ve sonra azalmaya başlar (C). Bu fazda bulunan tüm mikroplarda, bölünme birden durmaz. Bazıları yavaşta olsa, bu uygunsuz koşullara adapte olabilir ve üremelerini sürdürebilir. Ancak, böyle mikropların sayısı, tüm populasyona oranla çok azdır. Durma döneminin uzunluğu veya kısalığı, besi yerinin miktarı, bileşimi, üreyen mikrobun türü ve çevresel koşullara göre değişiklik gösterebilir. S. pneumoniae 'de çok kısa olun bu dönem, E.coli 'de 2-3 gün veya daha fazla olabilmektedir. Durma dönemi uzadıkça mikroorganizmaların morfolojilerinde (involusyon formları), kültürel özelliklerinde (R-formlarının artması) ve fizyolojik karakterlerinde (boyama reaksiyonları, pigment oluşumu, vs.) bazı değişmeler meydana gelir.
D) Ölme dönemi: Durma devresi değişmedikçe, mikroplar bu uygun olmayan koşullar altında ölmeye başlarlar. Populasyon, sayıca giderek, azalır. Ancak, bütün mikroplar ölmeyebilirler. Bazıları canlılıklarını koruyabilir. Bu nedenle de canlılık eğrisi sıfıra ulaşmaz (D). Kültürün steril hale gelmesi mikroplar arasında farklı bir durum gösterir. Örn. S. pneumoniae ve N. gonorrhoea iki üç gün içinde ölür ve otolize olur. E. coli 'de bu süre daha uzun ve M. tuberculosis 'de ise çok daha uzundur (aylar). Ölme döneminde canlı kalabilen mikropların morfolojilerinde değişiklikler meydana gelir (involusyon formları).
Sıvı ortamlarda mikropların ölmesi, aynen ısı ve ultraviolet ışınları etkisi ile ölmede olduğu gibi logaritmik bir tarzda oluşur ve aşağıdaki formül yardımı ile hesaplanır.
(S=S0e-kt )
S0 = Sıfır zamanında canlı mikrop sayısı
S = Belli bir süre (t) sonra canlı mikrop sayısı,
e = Doğal logaritma (2.7182)
k = Ölüm sabitesi
Eşit zaman aralıklarında populasyonun yarısı ölür. Örn. bir kültür de 100 milyon mikrop varsa, belli süre aralıklarından sonra, 50 milyon, 25 milyon, vs. canlı hücre kalır.
Senkron üreme: Saf kültürlerde bulunan mikropların hepsi aynı anda bölünmezler. Bir kısmı bölünürken bir kısmı olgunlaşma dönemindedirler. Ancak, kültürlerde bulunan mikropların, kısa bir süre için, bölünmelerine mani olunabilir veya aynı anda bölünmeleri sağlanabilir (senkron üreme). Bunu sağlamak için bazı yöntemler geliştirilmiştir. Örn. E. coli, timin (DNA'nın bileşiminde bulunan nukleotid bazı) olmayan ortamlarda bölünemezler. Eğer besi yerine bu madde katılırsa, mikropların çoğu aynı anda bölünmeye başlarlar. Bazen de mikroplar düşük sıcaklıkta (15 °C' de) 15-20 dakika tutulduktan sonra hemen optimal ısıya (37 °C' de) bırakılırsa yine aynı tarzda senkron üreme elde edilebilir. Senkron üreme zaman aralıklarıyla uygulanırsa, üreme eğrisi, merdiven basamakları gibi bir durum alır. Filtrasyon yöntemiyle de senkron üreme elde edilebilir.
03. Katı Ortamlarda Üreme
Katı ortamda ekilen mikroorganizmalar, sıvı besi yerlerine oranla daha sınırlı bir üreme şansına sahiptirler. Koloniler oluşmaya başladıktan sonra birçok faktörlerin etkisi altına girerler ve bunlar da büyüme ve üreme üzerine genellikle olumsuz yönde etkilerler. Bu nedenlerin başında;
1- Kolonilerin üst kısmında veya tepe bölgesinde bulunan mikroplara gıda maddelerin diffusyonla ulaşması güçleşir.
2- Kolonilerin altında ve civarında bulunan gıda maddeleri kısa bir süre sonra azalmaya başlar ve buna bağlı olarak da diffusyon sınırlanır.
3- Kolonilerden uzak bölgelerde bulunan gıda maddelerinin koloniye kadar gelebilme olasılığı azalır.
4- Hücre içinde oluşan metabolizma artıkları ve toksik maddeler dışarı atılamaz ve hücre içinde birikirler. Bu nedenle de metabolizma bozulur ve üreme kısıtlanır. Aynı durum, hücre dışına çıkması gereken ve çok lüzumlu göreve sahip olan enzimler için de söz konusudur.
5- Kolonilerin en üstündeki mikroorganizmalar gıdayı yeterince alamadıkları için dejenere olmaya başlarlar.
6- Etrafta gelişen diğer koloniler nedeniyle, beslenme alanı giderek daralır.
7- Basınç nedeni ile koloninin alt yüzünde bulunan mikroplar da dejeneresyon görülebilir. Ancak petri kutusu etüve ters konulduğundan bu basınç üreme döneminde oldukça azdır.
8- En alttaki mikroplardan dışarı çıkabilen toksik maddeler agarın yüzeyine yayılırlar.
9- Mikroplar birbirine bitişik durumda olduklarında, üreme kısıtlanır. Ancak, kenardakiler serbest üreme şansına sahiptirler.
10- Katı ortamlarda gıda diffüsyonu genellikle güçtür.
11- İnkubasyon ısısı nedeniyle, besi yerinden devamlı su kaybı, ortamın kurumasına ve diffusyonun daha zorlaşmasına sebep olur.
Yukarıda bildirilen nedenlerle bir bakterinin katı ortamda üremesi, sıvı besi yerlerine oranla daha zor olmakta, generasyon süresi, sayısı ve populasyon miktarı bakımından da noksanlıklar göstermektedir. Katı ortamlar ne kadar yumuşak ve koloni sayısı da az olursa, gelişme ve üreme daha fazla olur ve koloni çapı büyür.
Kolonilerin uygun besi yerlerinde gözle görülebilir bir düzeye ulaşabilmesi için geçen süre mikroorganizmalar arasında değişiklikler gösterir. Bu durum, mikropların cins ve türlerine özgü bir karakterdedir. E. coli, P. vulgaris, P. aeruginosa, S. aureus ve diğer bazı mikroplar 24 saat içinde veya daha kısa bir sürede gözle görülebilecek büyüklükte koloniler oluşturmasına karşın, S. enteritidis, S. pyogenes, C. pyogenes genellikle 48 saatten sonra, B. abortus ve diğer brusella türleri, PPLO'lar 4-5 günden sonra görülebilir düzeyde koloniler meydana getirirler. M. tuberculosis 'de bu süre 15-20 gün kadar sürer.
Koloni içinde mikroorganizmalar yığınlar halinde bulunurlar. Bu nedenle streptokok ve stafilokokların birbirlerinden ayırmada, katı besi yerinden ziyade sıvı ortamlarda üreyen kültürlerden boyalı preparatlar hazırlanmalıdır. Katı ortamlardan yapılan boyamalarda streptokoklar, stafilokok gibi görülebilirler.
Kolonilerin rengi, kokusu, yapısı, şekli, büyüklüğü, parlaklığı, besi yerine yapışma durumu, hemoliz yapma özelliği, ve diğer karakterleri mikroorganizmalar arasında farklar gösterir ve bu özellikler, cinslere, türlere, besi yerlerine, çevresel koşullara bağlıdır.
04. Üremenin Ölçülmesi
Mikroorganizmaların üremelerini saptamak için uygun optimal koşullarda üremiş kültürler kullanılmalıdır. Belli bir süre inkübatörde tutulduktan sonra mikropların üremesi çeşitli yöntemlerle tayin edilir.
1- Hacmin ölçülmesi: Belli bir süre sıvı besiyeri içinde üreyen mikroplar, dibi konik ve taksimatlı santrifüj tüplerine alınarak kuvvetlice santrifüje edilir. Üsteki sıvı atıldıktan sonra, dipte kalan tortunun miktarı santimetreküp olarak değerlendirilir ve hacim olarak ölçüsü bulunur.
2- Kuru ağırlık tayini: Santrifüj yardımı ile çöktürülen ve distile su ile birkaç kez yıkanan belli miktardaki kültürde bulunan mikroorganizmalar, ısıya dayanıklı belli ağırlıktaki kaba konarak 120°C. 3 saat ısıtılır. Soğuduktan sonra tartılır. Ağırlığı sabit kalıncaya kadar ısıtma ve soğutma işlemine devam edilir. Böylece elde edilen ve değişmeyen ağırlık, mikropların kuru ağırlığını verir. Kuru ağırlığın miktarı, üremenin çokluğu veya populasyonun miktarı ile ilişkilidir.
3- Yaş ağırlık tayini: Santrifüjle çöktürülen ve birkaç kez yıkanan tortu, ya tüple birlikte veya ayrı olarak tartılır. Bu suretle belli bir süre ve miktardaki kültürdeki bakteri ağırlığı saptanmış olur.
4- Total azot tayini: Bakteri içinde bulunan maddelerdeki nitrogen miktarının tayini duyarlı yöntemler arasında bulunmaktadır. Bu tür nitrogen miktarının bulunmasında mikro-Kjeldahl tekniğinden yararlanılır.
5- Bulanıklık (türbidite) tayini: Belli miktardaki kültürlerde muayyen süre inkube edilmiş olan mikropların üreme durumu, ya standart bulanıklık tüpleri (McFarland, Brown) ile karşılaştırarak veya spektrofotometrik yöntemle (optikal densite tayini ile) tesbit edilir.
6- Sayma suretiyle: Katı ortamlarda üreme durumunu anlamak için birçok yöntemler uygulanırsa da en çok kullanılanı koloni sayma tekniğidir.
Başlıca mikrop sayma teknikleri:
a) Boyalı preparatlarda, mikroskop altında direk sayma tekniği,
b) Petroff-Hause aleti ile mikrop sayma yöntemi,
c) Breed metodu ile frotideki mikropların sayılması,
d) Karşılaştırma tekniği ile mikropların miktarının bulunması,
e) Partikül sayımı (Coulter counter ve diğer aletlerle),
f) Dilusyonla sayma tekniği,
g) Koloni sayma usulü,
h) Sellüloz filtreleri ile mikrop sayımı
1 Kaynak: Temel Mikrobiyoloji