Transkripsiyon (mRNA Sentezi)
Prof. Dr. Mustafa Arda
Ankara Üniversitesi Veteriner Fakültesi
Temel Mikrobiyoloji 1
01. Genel Giriş
02. Başlama
03. Zincir Uzaması
04. Terminasyon (Bitiş)
01.Genel Giriş
Protein sentezinde başlıca 3 tür RNA'nın etkinliği vardır. Bunlar da mRNA (mesenger RNA), tRNA (transfer RNA) ve rRNA (ribosomal RNA)'dır. Bunların dışında, DNA'nın replikasyonunda geçici olarak görev yapan ve primase enziminin katalitik etkisi ile sentezlenen 15-20 nukleotid'den oluşan primer RNA (pRNA) da bulunmaktadır.
Mesenger RNA (mRNA), DNA iplikçiliklerinin birinden (3' ¬ '5) yönünde olanından, kalıp DNA, sens DNA), RNA polimerase enziminin (transkriptase) katalitik etkisiyle sentezlenen (transkripsiyon) ve amino asit veya polipeptid sentezi için gerekli bilgileri kodlar halinde taşıyan, tek iplikçikli bir nukleotid sekansıdır.
DNA'nın bir iplikçiğinde bulunan genetik bilgilerin şifreler (kodlar), halinde tek iplikçik RNA'ya transferine transkripsiyon ve bu bilgileri taşıyan RNA'ya da mesenger RNA adı verilir.
Prokaryotik mRNA çok dayanıksızdır ve translasyon başladıktan hemen sonra 5' -ucundan metabolize olmaya başlar. Ortalama ömrü 1-3 dk. kadardır. Ökaryotik mRNA ise saatlerce dayanır.
Prokaryotik mRNA, birden fazla genin (proteinin) kodlarına sahiptir (polisistronik mRNA veya poligenik mRNA). Buna karşın ökaryotiklerin mRNA'sı monosistronik (monogenik) bir özellik taşır.
Transfer RNA (tRNA), protein sentezinde, sentetase enzimi tarafından aktive edilen amino asitlere bağlanarak bunları, ribosomlar üzerinde sıraya giren kodon'larına taşıyan adaptör bir moleküldür. Yaklaşık 75-95 nukleotidden oluşur ve fonksiyonuna göre 5 kollu bir sekonder yapıya sahiptir. Bakteri hücresinde yaklaşık 50 farklı tRNA bulunur. Her amino asit için bir veya birden fazla tRNA vardır ve spesifik sentetase enziminin katalitik etkisiyle, her amino asit, kendine ait tRNA'nın 3' -CCA ucuna bağlanır. Bu enzim kendine ait hem amino asiti ve hem de tRNA'yı tanır. Transfer RNA üç boyutlu (primer, sekonder, tersiyer) bir yapıya ve 5 kola (D-kolu, antikodon kolu, ekstrakol, T y C kolu Akseptör kol,) sahiptir.
Ribosomal RNA (r RNA), ribosomun yapısal ve fonksiyonel bir komponentidir. Prokaryotiklerde 70 S ribosom 2 alt üniteden oluşur. Bunlardan biri 50 S alt ünite (23 S rRNA, 5S rRNA ve proteinler) ve 30 S alt ünite (16S rRNA ve proteinler) dir.
Ribosomlar spontan olarak asamble olurlar.
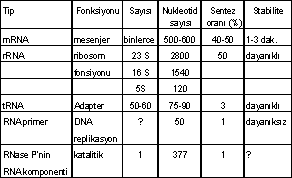
Bu 3 RNA için de, genetik bilgileri şifreler halinde taşıyan ve bu nedenle de, DNA gibi, genetik bir karakter gösteren mesenger RNA (mRNA)'dır.
Prokaryotiklerde, transkripsiyon ve translasyon sitoplazmada gerçekleşmesine karşın, ökaryotiklerde gerek primer mRNA ve gerekse olgun mRNA'nın sentezi çekirdekte tamamlanır ve olgun mRNA buradan sitoplazmaya geçerek transle edilir. Primer mRNA'larda, ekzonlarla birlikte intronlarda bulunmasına karşın, olgun mRNA'da sadece ekzonlar vardır intronlar çıkarılmıştır.
Bakterilerde, çift iplikçikli, sarmal ve sirküler olan DNA molekülünden, 3' ® 5' yönündeki tek iplikçiğinde bulunan genetik bilgilerin kodlar halinde, RNA polimerase enziminin katalitik etkisiyle, yeni sentezlenen tek iplikçik mRNA'ya (mesenger RNA) aktarılmasına transkripsiyon adı verilir. Mesenger RNA, protein zincirindeki amino asitlerin türlerini ve sıralarını, nukleotid sekansları halinde (genetik mesajlar halinde) taşır. Ancak, DNA'daki Timin yerine, mRNA'da Urasil bulunur. Sentezlenen mRNA 5' ® 3' yönünde olup kendisine kalıp ödevi gören DNA iplikçiğine (3' ® 5') de, anti paraleldir.
Sentez işlemini gerçekleştiren RNA polimerase enzimi (transkriptase, DNA'ya bağımlı RNA polimerase) elektroforetik analizde 5 farklı alt üniteye ayrılmaktadır. Bunlar da; 2 molekül halinde a2 (36 500), iki ayrı b, b', (151 000 ve 155 000) ve bir de sigma faktörüdür (s, 70.000). Toplam enzim 480 000 M.A.'dadır. Alt proteinlerden ilk 4 üne ( a2, b, b') merkez enzim kompleksi adı verilir. Bunlardan a faktörleri mRNA zincirinin başlamasında, b-'faktörü başlama ve zincirin uzamasında, b' faktörü DNA'ya bağlanmada ve sigma faktörü de enzim tarafından promotorun tanınmasında etkin rollere sahiptirler. Sigma faktörü, sentez başladıktan sonra hemen ayrılarak sitosola döner ve sentezi merkez enzim kompleksi devam ettirir. E. coli 'de sigma faktörünün, tek olduğu bildirilmişse de(s70), bunun dışında d32, d54, d24 gibi varyantlarının da var olduğu sigma faktörü olmadan merkez enzimleri promotora sağlamca bağlanamazlar.
Prokaryotiklerde (bakterilerde) transkripsiyon başlıca 3 aşamada gerçekleşmektedir.
1) Başlama
2) Zincir uzaması
3) Bitiş
02. Başlama
Mesenger RNA'nın sentezinin başlangıcında, RNA polimerase enzimi, DNA'nın 5' -ucuna doğru, primidinlerden zengin ve promotor olarak adlandırılan bölgesine bağlanır. Bu bağlanma 77-80 bazlık bir bölgeyi (-55 ile +20 bazları arası) kapsar. Böylece, aynı zamanda, yaklaşık 60 bazlık bölge RNase'ların etkisinden de korunmuş olur (Burada (-) olarak ifade edilen bölge, mRNA'da başlangıç kodonunu (AUG) oluşturan sıfır noktasından (başlama noktası) 5' -ucuna doğru (sola doğru, upstream), (+) ise 3' -ucuna doğru (sağa doğru, downstream) olan bölgeleri ifade eder). Bu -55 bölgesine kadar olan bölge içinde çok korunmuş olan ve merkezleri -10 ile -35'e isabet eden, her biri yaklaşık 6 bazdan oluşan iki bölge bulunmaktadır. Bunlardan birincisine Pribnow box adı da verilmekte, burası -6 ile -13 bölgeleri arasında yer almakta ve TATAAT bazlarından oluşmaktadır. İkinci bölge ise -30. ile -37. bazlar arasında bulunmakta ve T T G A C A bazlarından meydana gelmiştir. Bu kısımlara başlama bölgeleri (start bölgeleri) adı da verilmektedir. Bu bölgelerden -10, DNA'da açılma sinyalini ve -35'de RNA polimerazın bölgeyi tanıma sinyalini taşırlar.
E. coli ‘deki promotor bölgesi yandaki şekilde gösterilmektedir. | 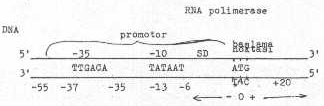
|
Başlama noktası (start point) ile -10 bölgesi arasında bulunan ve yaklaşık 6-7 bazdan oluşan kısım Shine Dalgarno (SD) sekansları olarak tanımlanır ve burası da aynen mRNA ya aktarılır. Buna karşın, -10 ve -35'lik bölgeler (promotor bölgesi) sentezin başlamasında rol aldığı halde, mRNA'ya transfer edilmezler.
mRNA'nın sentezinin gerçekleştiği DNA'nın 3' ® 5' yönündeki iplikçiğine kalıp sekans (anlamlı iplikçik) ve diğer iplikçiğe (5' ® 3') ise kodlayan sekans (kodlayan iplikçik) adı verilir. Sentezlenen mRNA'da 5' ® 3' yönünde olup, kendine kalıplık görevi yapan DNA iplikçiğine (3'®5'), anti paraleldir. Diğer bir deyimle mRNA, DNA'daki, kodlayan sekansa identik, kalıp DNA iplikçiğine de anti paraleldir.
Bakterilerde transformasyonun aşamaları ve sonlandırılması yandaki şekilde gösterilmektedir. | 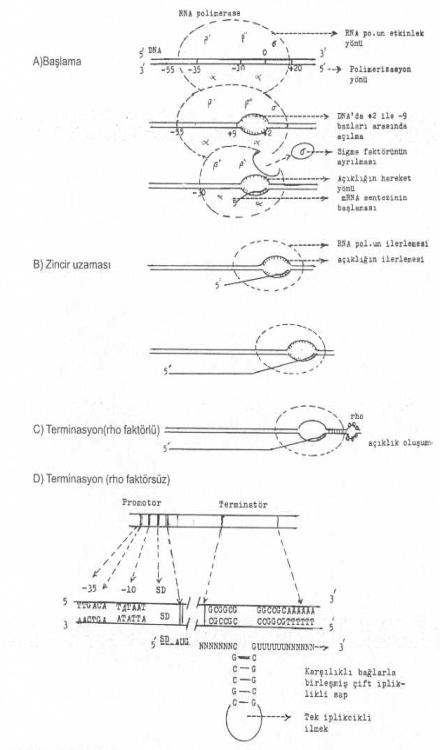
|
DNA'da -10 ile -35 bölgesinde oluşan mutasyonlar, RNA polimerase enziminin bağlanmasına ve polimerizasyona olumsuz yönde etkiler.
Bazı promotorlarda, yukarda belirtilen ve çok korunan, -10 ve -35 bölgelerinin yerleri, promotorun kuvvetine göre değişebilir ve böylece promoterler etkinliklerine göre zayıf, orta ve kuvvetli promoterler olmak üzere gruplandırılabilirler (promotorun etkinliği, belli bir süre içinde sentezlenen proteinlerin miktarına göre, tanımlanmaktadır). Bazı promotorlar bir genin sentezini bir kaç saniyede başlatabilir ve bazıları da birkaç dakikada gerçekleştirebilecek durumdadır. Buna bağlı olarak da hücre içinde bazı proteinler az ve bazıları da fazla bulunmaktadırlar.
RNA polimerazın bağlantısı, bu iki bölgede kuvvetlendikten (yaklaşık 10 saniyede) sonra, promotorda hemen açılma olmaz. Önce kısa bir süre için kapalı kalır ve bunun hemen arkasından, -9 ile -2 sekansları arasında yaklaşık 10 baz uzunlukta DNA'nın 5' ® 3' yönündeki iplikçiğinde, bir açılma meydana gelir. Bu bölgede, DNA'da karşılıklı bazlar arası bağlantılarda kopmalar oluşur. Bu açılmayı RNA polimerase enzimi gerçekleştirir. Açılma meydana geldikten hemen sonra da, DNA'nın 3' ® 5' yönündeki iplikçiğinden ve başlangıç noktasına yakın yerinden sentez başlar. Sentez başlayınca da sigma faktörü kompleksten ayrılır. Ayrılma olunca, merkez enzimleri (a2, b, b') polimerizasyona deva ederler. Ancak, RNA polimerazın DNA üzerindeki kapladığı bölge 77 bazdan 50 bazlık bir alana iner (-30 ile +20). Sigma faktörü, RNA polimerase enziminin DNA'nın 5' -ucuna bağlanmasını güvence altına alır.
Sentezi, terminasyon bölgesine kadar merkez enzimleri (a2, b, b') sürdürür. Sigma faktörü, sentezin başlangıcından enzim promotora sağlam ve güvenle yerleştikten sonra (10 baz sentezlendikten sonra) ayrılmaktadır. Sigma faktörü olmazsa enzim başka yerlere de bağlanabilir. Bu nedenle sigma faktörü, enzimin promotora ve doğru yerlere bağlanmasını garanti altına alır.
03. Zincir Uzaması
Polimeraz enzimi tarafından, promotor bölgesinde -9 ile +2 bazları arasında (yaklaşık 10-11 baz uzunlukta) bir açıklık meydana getirildikten hemen sonra da (2-10 saniye içinde) mRNA sentezi de başlar. Açılma ile birlikte RNA pol. enzimi, DNA üzerinde, 5' ® 3' yönünde ilerlemeye başlarken, açık olan kısmı da, DNA'nın 3' ® 5' yönündeki iplikçiğin üzerinde de aynı yönde mRNA sentezlemeye devam eder.
Açılan DNA bölgesinin hemen arkasından da ilmek kısmında kapanma meydana gelir. Açılma veya açıklık aynen, enzimle birlikte ilerler. Böylece, DNA'nın dejenerasyonu önlenmiş olur.
Yeni sentezlenen mRNA ancak geçici olarak (kısa bir süre için) üç bazla, DNA'ya bağlanmıştır (DNAxRNA hibrid dubleks molekül). Başlangıçta, önce 8-9 bazlık kısa bir sekans sentezlenerek hemen çıkarılırlar. Çıkan bu kısımlarla, RNA pol.un doğru yere bağlandığı ve sentezin doğru yönde olduğu güven altına alınır. Bundan sonra sentez 5' ® 3' yönünde aynen ve hızla devam eder. Sentez sırasında nukleotid trifosfatlar, mRNA'nın 3' -OH ucuna ilave edilerek tek iplikçik RNA'nın 5' ® 3' yönünde uzamasını sağlar. Ancak, DNA'da bulunan timin yerine, RNA'da urasil bulunur.
RNA polimerase, DNA üzerinde, hızla, DNA'yı açarak ilerlerken, arkadan da kapatır ve bir taraftan da mRNA zinciri uzamaya devam eder.
Bakterilerde RNA polimerase enziminin boyutları 90x95 x 160 A°, mayalarda 140x136x110 A° arasında olduğu belirtilmektedir. Enzimin yüzeyinde de 16 A° kadar bir kanalın bulunduğu ve bunun DNA boyunca ilerlediği de bildirilmektedir.
RNA zinciri 5' ® 3' yönünde uzarken, 37°C'de her saniyede yaklaşık 40 nukleotid zincire katılır (DNA replikasyonunda ise saniyede 800 bp DNA'ya ilave edilir). Bu durum mRNA zincirine, saniyede, yaklaşık 13 amino asitin kodonunun katıldığını ifade eder. İnsektlerde bu hız 20 nukleotid/sn'dir.
Bir bakteri içinde yaklaşık 7000 molekül kadar RNA pol. enzimi bulunduğu açıklanmıştır. Ancak, bunlardan 2000-3000 tanesi RNA sentezinde aktif olarak görev alır.
Hücre içinde ilk mRNA sentezi 2.5 dakika da ve bundan 30 sn. sonra da translasyon ve ilk protein sentezi başlar. Translasyon da yaklaşık 3 dk. devam eder. Buna göre 10.000 bp.'lik bir DNA segmentinin transkripsiyonu için 4-5 dakika yeterli olmaktadır. Bakterilerde, mRNA sentezi devam ederken, ilk önce sentezlenen 5' -ucu, ribosomlar da transle olmaya başlar. Böylece, transkripsiyon devam ederken translasyon da başlamış olur. Mesenger RNA, translasyon tamamlanınca 5' -ucundan itibaren degradasyona başlar. Ancak, ayrışma hızı, translasyondan daha yavaştır. Ökaryotiklerde mRNA'nın hücre içindeki dayanma süresi, prokaryotiklerden daha uzundur (4-24 saat).
Transkripsiyon, translasyon ve degradasyonun (mRNA'nın ayrışması) yönü, 5' ® 3'dür.
04. Terminasyon (Bitiş)
RNA polimerase, mRNA sentezini başlattıktan sonra zincir uzaması terminatör sekansa gelinceye kadar devam eder ve durur. Artık, enzim, zincire herhangi bir nukleotid ilave etmez. Sonra, zincir serbest kalır ve DNA'dan ayrılır. İlmekteki DNA x RNA dubleks hibrid bağları kopar. DNA'daki açıklık kapanır ve eski kapalı formuna döner.
Başlıca 2 tür terminasyon mekanizması bildirilmiştir.
1) Terminatör faktöre (rho faktörü) bağımsız sonlanma: Bu tür terminasyon da, RNA pol. enzimi (merkez enzimi) terminatör bölgeye ulaştığında sentez durur. Terminatör bölge iki yapısal karakter gösterir. Bunlardan biri G-C bazlarından zengin karşılıklı birleşmiş bir sap ve bunun ucunda tek iplikçik ilmek ile bu sapın tabanında 6 urudin bazından oluşan terminatör sekans bölgesi, diğeri ise a 6 adet A bazından (poli A) meydana gelmektedir.
RNA pol., G-C'den zengin bölgeye gelince yavaşlar ve durur. Çünkü, bu bazlar arasındaki bağlar oldukça sağlam ve stabildir ve açılmaları zordur. DNA'nın bu yapısı, oluşan mRNA'ya saç tokası gibi bir şekil de verdirir (Rho faktörsüz).
2) Terminatör faktöre (rho) bağımlı sonlanma: Bu tarz terminasyonda rho faktörü (M.A.: 46.000, hekzamer) RNA üzerinde özel bir yere sıkıca bağlanarak ribonukleotid trifosfatları, nukleotid difosfat halinde hidrolize eder. Burada oluşan enerji, Rho faktörünü mRNA üzerinde, DNA'daki açıklığa doğru iter ve burada durur. Açıklık RNA pol.'un daha ileri gitmesini önler ve RNA'nın serbest kalmasına yardımcı olur. Bundan sonra RNA pol., mRNA ve rho faktörü serbest kalırlar. Diğer bir terminasyon faktörü olan nus A gen ürünü protein de (asidik, M.A. 55 000), zincirinin serbest kalmasına katkıda bulunur.
Bazı durumlarda sentez terminasyon bölgesini de geçebilir, daha ileri gidebilir ve transkripsiyon bir süre daha devam edebilir. Başka bir terminasyon sekansına ulaşınca sentez durur.
[1] Kaynak : Temel Mikrobiyoloji